
by Jim Steele
Is bleaching the legacy of a marvelous adaptation mechanism or a prelude to extirpation?
A Warm Evolutionary Legacy
Despite increasing confirmation of the Adaptive Bleaching Hypothesis and its ability to explain coral resilience, most people are unaware of its debate within the scientific community. The ability to rapidly adjust to changing environments by modifying their symbiotic partnerships has been the key to their success for millions of years. As one expert wrote, the “flexibility in coral–algal symbiosis is likely to be a principal factor underlying the evolutionary success of these organisms”.
Our modern day reef-building corals first evolved in exceedingly warm and stable climates when deep ocean temperatures were 10°C higher than today and palm trees dotted the Antarctic coast. As ice caps began to form in Antarctica ~35 million years ago sea levels fell and warm epi‑continental seas dried. After ocean depths had cooled for another 30 million years, Arctic ice caps began to form and the earth entered an age with multiple episodes of glacier advances and retreats causing sea levels to rise and fall. Just eighteen thousand years ago during the last glacial maximum, all our shallow reefs did not exist, as sea levels were 400 feet lower than today.
The 35 million year cooling trend increasingly restricted reef-building corals to more tropical latitudes where winter water temperatures remain above 16 to 18 °C. As their evolutionary history would predict, today’s greatest concentrations and greatest diversity of corals are found in the earth’s persistently warmer waters, like the Indo-Pacific Warm Pool. Likewise species inhabiting our warmest waters have undergone the fewest episodes of severe coral bleaching. Given their evolutionary history, coral’s greatest achievement has been enduring bouts of sustained climate cooling and rapid temperature swings. Even during warm interglacials coral battled cold temperatures dips. Studies of 7000-year-old fossil coral reefs in the South China Sea revealed high coral mortality every 50 years due to winter cooling events. Indeed most researchers believe past coral extinctions were most commonly due to cold events. Accordingly research has estimated that during the cold nadir of each ice age, coral reef extent was reduced by 80% and carbonate production was reduced by 73% relative to today.
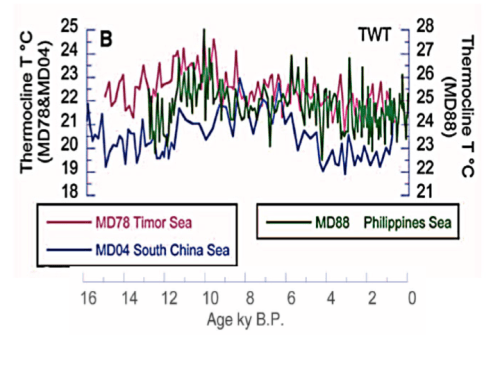 As the last ice age ended, coral expanded their range with warming temperatures. At the peak of the Holocene Optimum 10,000 years BP (Before Present), coral adapted to tropical ocean temperatures in the heart of the Coral Triangle were 2.1 °C warmer than today. As illustrated above, temperatures cooled since then but frequently spiked or plummeted by 2 to 3 degrees over the course of a few centuries. One thousand years ago during the Medieval Warm Period, coral thrived in Pacific water masses that were ~0.65° warmer than in recent decades, then cooled ~0.9°C by the 1700s. Given coral’s evolutionary history, it is unlikely coral were better adapted to 1800s Little Ice Age temperatures versus Medieval Warm Period or 20th century temperatures. Emerging research now suggests coral bleaching has been an integral part of corals’ adjustment mechanisms to an ever-changing environment.
As the last ice age ended, coral expanded their range with warming temperatures. At the peak of the Holocene Optimum 10,000 years BP (Before Present), coral adapted to tropical ocean temperatures in the heart of the Coral Triangle were 2.1 °C warmer than today. As illustrated above, temperatures cooled since then but frequently spiked or plummeted by 2 to 3 degrees over the course of a few centuries. One thousand years ago during the Medieval Warm Period, coral thrived in Pacific water masses that were ~0.65° warmer than in recent decades, then cooled ~0.9°C by the 1700s. Given coral’s evolutionary history, it is unlikely coral were better adapted to 1800s Little Ice Age temperatures versus Medieval Warm Period or 20th century temperatures. Emerging research now suggests coral bleaching has been an integral part of corals’ adjustment mechanisms to an ever-changing environment.
Coral Mortality and Resilience
There are 4 widespread misconceptions about bleaching propagated by tabloid media hyping climate doom and researchers like Hoegh-Guldberg. To clarify:
- Bleaching is not always driven by warming temperatures
- Bleaching is not responsible for most coral mortality.
- Coral can rapidly respond to disturbances and replace lost cover within a decade or less.
- Bleaching, whether or not it results in coral mortality, is part of a natural selection process from which better-adapted populations emerge.
Multiple causes of bleaching
In contrast to researchers like Hoegh-Guldberg who emphasizes coral bleaching as a deadly product of global warming, bleaching is a visible stage in a complex set of acclimation mechanisms during which coral expel, shift and shuffle their symbionts, seeking the most beneficial partnership possible. Bleaching can be induced by stressful interactions between temperatures, disease, heavy rains, high irradiance from clear skies and competition with seaweeds. Indeed abrupt warm water events like El Nino have induced widespread bleaching and high mortality. But or of colder waters have also induced bleaching.In contrast to researchers like Hoegh-Guldberg who emphasizes coral bleaching as a deadly product of global warming, bleaching is a visible stage in a complex set of acclimation mechanisms during which coral expel, shift and shuffle their symbionts, seeking the most beneficial partnership possible. Bleaching can be induced by stressful interactions between temperatures, disease, heavy rains, high irradiance from clear skies and competition with seaweeds. Indeed abrupt warm water events like El Nino have induced widespread bleaching and high mortality. But cold winters or La Nina induced upwelling of colder waters have also induced bleaching.
NOAA has also contributed to these misconceptions by overemphasizing just warm-event bleaching. On NOAA‘s web page “What is Coral Bleaching”, NOAA reported, “the U.S. lost half of its coral reefs in the Caribbean” in one year due to warmer waters. But the Caribbean’s main cause of lost reefs was due to an outbreak of the White Band disease in 1981-82. White band specifically targets members of the genus Acropora, like the Staghorn and Elkhorn coral, reducing by 80% of their cover that once dominated the Caribbean reefs. However since the mid 80s experts reported coral cover has changed relatively little.
NOAA also downplayed cold temperature bleaching stating the 2010 cold event just “resulted in some coral death.” However NOAA’s statement stands in stark contrast to coral experts who reported the January 2010 cold snap was the worst coral bleaching and mortality event on record for Florida’s Reef Tract. They reported, “the mean percent coral mortality recorded for all species and subregions was 11.5% in the 2010 winter, compared to 0.5% recorded in the previous five summers, including years like 2005 where warm-water bleaching was prevalent.” Globally there has been an increase in observed cold bleaching events and 2010 was Florida’s first cold bleaching since the 1970s. Globally there have been several more reports of cold induced bleaching and then recovery as the waters warmed.
There is a perception that bleaching suddenly became more common only since the 1980s, leading some to speculate bleaching is due to rising CO2 and global warming. However, whether warming since the Little Ice Age is natural or anthropogenic, warming does not explain the increased observations of cold bleaching. More frequent observations of bleaching events may be partially due to the advent of remote sensing satellites that have allowed greater global coverage only since the 1980s. Furthermore determination of bleaching severity and mortality requires teams of divers to ground truth satellite data and fine-tune percentages of affected reefs. But SCUBA diving only became possible in the decades after Jacques Cousteau invented the Aqualung in the 1940s. Although natural rates of warming during the 30s and 40s were similar to today, coral reef studies were also hampered by the unsafe battleground between Japan and the Allies. War-time efforts such as the Battle of the Coral Sea, and fights to control the islands of Peleliu, Midway, Iwo Jima, the Philippines, or subsequent nuclear testing on the Bikini Atoll. The resulting reef devastation likely obscured any natural bleaching events.
We now know bleaching regularly happens due to seasonal fluctuations between high solar irradiance and warm temperatures of summer versus lower irradiance and cooler temperatures in winter. High irradiance can damage the corals’ symbiotic algae when photosynthesis runs too rapidly, while low irradiance detrimentally reduces photosynthetic output. Thus coral undergo natural adjustments to seasonal changes by expelling a portion of their symbiotic algae in summer. This leads to temporary or partial bleaching. Low light and colder temperatures slow photosynthesis, so coral increase their symbiont density in winter.
Similarly in response to changes in sunlight, the same species will alter their symbiotic partnerships as irradiance declines at increasing depths or when and where water turbidity alters irradiance. Bleaching is often temporary and mild as coral shuffle and switch their symbiotic algae in order to adapt, but sustained extremes, warm or cold, can prolong bleaching and starve the coral. Whether coral die or not depends on how quickly new symbionts are acquired relative to how much energy the coral has stored, or coral’s ability to feed on plankton as an alternative energy source.
All recent global bleaching events have been driven by El Nino events. The 1998 El Nino caused widespread mortality, an estimated 16% globally. Observed bleaching in response to warm tropical waters invading cooler regions aroused fears that climate change had contributed to this “unprecedented” event. However researchers have noted the relationship between warmer ocean temperatures and “bleaching has been equivocal and sometimes negative when the coolest regions were not in the analyses.” In other words coral living in the warmest waters were well acclimated to the warmest waters redistributed by an El Nino. Furthermore mortality did not always occur during periods with the warmest temperatures, but during the winter or ensuing cold La Nina conditions. Such observations suggest the rapid swings between anomalously warm El Nino and anomalously cold La Nina conditions are the most stressful.
Stressful rapid temperature variations due to El Nino events have occurred throughout the past 10,000 years. As illustrated below from Zhang 2014, the frequency of El Ninos during the past century has been neither extremely high, nor extremely low. Most living coral species have survived over a million years of climate change and have endured the extreme El Nino frequencies of the past 3000 years including the Little Ice Age. El Nino events are a function of natural ocean variability and there is no consensus regards any effect from rising CO2 on El Nino frequency or intensity. To survive extremes from past natural variability, coral species had to be extremely resilient in ways that are just now being understood.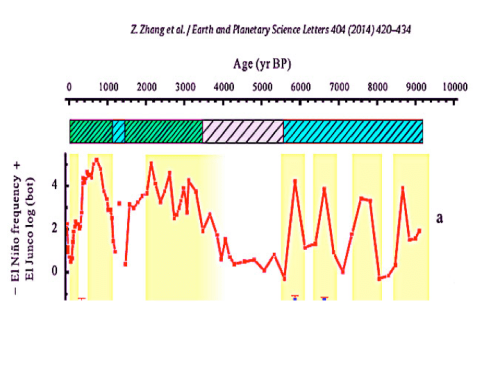 Bleaching Causes the Least Mortality
Bleaching Causes the Least Mortality
Most extreme bleaching events are associated with El Ninos, but the high mortality rates are not just a function of higher temperatures. Due to associated flooding and high rainfall, the resulting change in salinity disrupts coral osmosis, which can result in coral death. Furthermore tropical storms and heavy wave action are a major cause of lost coral reefs, but storms also bring heavy rains that also induce bleaching. Although some try to link storm-related mortality to climate change, there is no evidence of an increasing trend in tropical storms. As illustrated by the pie graph from Osborne 2011, in the Great Barrier Reef the explosion of the coral-eating Crown of Thorns starfish (A. planci) and tropical storms contributed to the greatest loss of coral colonies, 70.5%. Bleaching is a very minor contributor to coral mortality, just 5.6%, and that bleaching can be induced by warm or cold temperatures, heavy rains and floods or high irradiance from anomalously clear skies.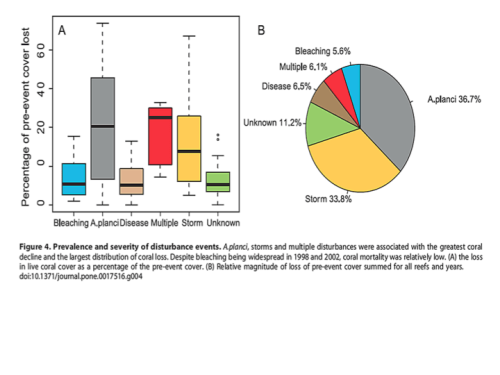 Due to coral’s symbiotic efficiency and recycling of nutrients, corals dominate in nutrient-limited tropical waters. Normally those low nutrient conditions also prevent predators like the Crown of Thorns starfish (COTS) from rapidly reproducing because their plankton-feeding larvae typically starve. But increased inflow of nutrients due to landscape changes, agriculture run-off and sewage, has increased plankton blooms and thus the survivorship of COTS’ larvae. The ensuing population explosions of coral eating adults have decimated many reefs. COTS does not exist in the Caribbean. Instead coral there are battling bacterial diseases like white-band that can be spread by coral-eating snails. Humans have indeed tipped the balance in favor of COTS and in addition to destructive over fishing with dynamite and cyanide, those causes of coral death are the only factors we can remedy.
Due to coral’s symbiotic efficiency and recycling of nutrients, corals dominate in nutrient-limited tropical waters. Normally those low nutrient conditions also prevent predators like the Crown of Thorns starfish (COTS) from rapidly reproducing because their plankton-feeding larvae typically starve. But increased inflow of nutrients due to landscape changes, agriculture run-off and sewage, has increased plankton blooms and thus the survivorship of COTS’ larvae. The ensuing population explosions of coral eating adults have decimated many reefs. COTS does not exist in the Caribbean. Instead coral there are battling bacterial diseases like white-band that can be spread by coral-eating snails. Humans have indeed tipped the balance in favor of COTS and in addition to destructive over fishing with dynamite and cyanide, those causes of coral death are the only factors we can remedy.
To understand coral resilience in the face of the variety of onslaughts, coral reefs must be seen as dynamic systems that oscillate over decadal periods, as well as centuries and millennia. Snapshots focused only on a few years when coral reefs decline misrepresents coral resilience and promotes false gloom and doom, as well as useless management plans. A long-term study of coral ecosystems of an island in French Polynesia demonstrates corals’ dynamics response to 32-years of storms, Crown of Thorns starfish and bleaching. Coral mortality is often measured as a function of the change in “coral cover”, and 45 to 50% of the healthy reef system around the island of Tiahura was covered with coral.
As illustrated below in Figure 1 from Lamy 2016, an outbreak of COTS removed 80% of the live coral cover between 1979 and 1982, reducing total coral cover to 10% of the reef. However by 1991 the coral had fully recovered. As designated by the small gray arrows at the top, three bleaching events occurred during that recovery period. Later destruction from a 1991 cyclone again reduced coral cover but again coral recovered reaching its greatest coverage of 50% by the year 2000. And again during that recovery there were 3 more bleaching events. Since 2006 the coral suffered their greatest loss due to another outbreak of COTS, quickly followed by another cyclone. High mortality promoted high seaweed cover (dotted green line) that has inhibited coral recovery. Over that time, coral bleaching was associated with periods of recovery, suggesting little if any detrimental effects. As will become clear shortly, one also could reasonably argue those bleaching events were beneficial..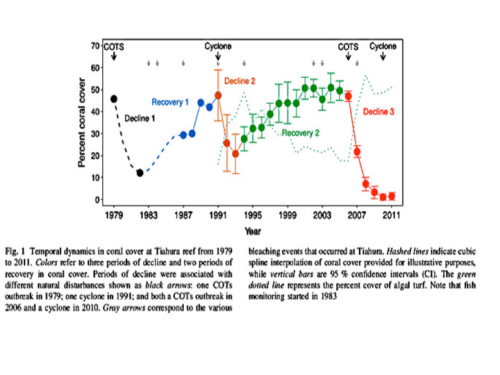
Rapid Coral Recovery:
Tiahura’s coral recovery periods typically required 7 to ten years, and appeared to be unaffected by the 1998 El Nino. Several other studies have reported similar recovery periods, but some locations required 10 to 20 years to fully recover. In Australia’s Great Barrier Reef (GBF), the 1998 El Nino induced above average sea surface temperatures and salinity changes for 2 months triggering massive coral losses in the reef’s upper 20 meters. At the GBF’s Scott Reef, the upper 3 meters lost 80 to 90% of its living coral and the disappearance of half of the coral genera. Yet researchers observed, “within 12 years coral cover, recruitment, generic diversity, and community structure were again similar to the pre-bleaching years.” A similar long-term study in the Maldives observed a dramatic loss of coral during the 1998 El Nino but by 2013 the reefs also had returned to “pre-bleaching values”. Although a reef’s recovery sometime requires re-colonization by larvae from other reefs, a process known as re-sheeting or Phoenix effect can facilitate a reef’s speedy recovery. Often a small percentage of living “cryptic” polyps with a more resilient symbiotic partnership were embedded within a “dead” colony and survive extreme bleaching. They then multiply and rapidly “re-sheet” the colony’s skeletal remains.
In addition to rapid recovery of coral cover, researchers are finding bleached reefs have been increasingly less susceptible to subsequent bleaching. For example studies in Indonesian waters determined that two coral species, highly susceptible to bleaching, had experienced 94% and 87% colony deaths during the 1998 El Nino. Yet those same species were among the least susceptible to bleaching in the 2010 El Nino, with only 5% and 12% colony deaths despite a similar increase in water temperatures. Similarly, changes in resilience were observed in response to cold water bleaching in the Gulf of California. Increased resilience in response to a variety of bleaching events prompted the Adaptive Bleaching Hypothesis first proposed in 1993. The hypothesis suggests that although bleaching events are a response to stress, it creates the potential for coral to acquire totally new and different symbionts that are better suited to those stressful conditions. Contrary to Hoegh-Guldberg’s claim that coral reef systems will “experience near annual bleaching events that exceed the extent of the 1998 bleaching event by the year 2040”, scientists are increasingly observing the exact opposite. After reefs recover from severe bleaching, colonies have evolved enhanced resilience to future bleaching.
Coral Symbiosis, Symbiont Shuffling and Rapid Adaptation
 A single coral colony is comprised of 100s to millions of individual “polyps” (seen above). Each polyp can be visualized as an upside down jellyfish (coral’s close cousins) with their backs cemented to a surface and tentacles extended outward to capture passing food particles, live prey, or new symbionts. However because coral live in nutrient depleted environments, in addition to filter feeding, polyps harbor single-celled photosynthesizing symbionts inside their cells. Those symbionts (aka zooxanthellae) typically provide ~90% of the coral’s energy needs. Just 40 years ago it was believed all corals were host to just one photosynthesizing symbiont, a single species from the dinoflagellate genus Symbiodinium. But thanks to technological advances in genetic sequencing, we now know a coral species can harbor several potential species or types of Symbiodinium algae, each capable of responding optimally to a different set of environmental conditions and coral physiology. As predicted by the adaptive bleaching hypothesis, improved genetic techniques have revealed a wondrously diverse community of symbionts that coral can choose from. Coral can no longer be viewed as organisms that only adapt slowly over evolutionary millennia via genetic mutation and natural selection. Coral must be seen as an “eco-species” (aka holobiont) that emerges from the synergy of the coral and its varied symbionts. And we now know those emergent eco-species can rapidly evolve with changing climates by shuffling and shifting those symbionts.
A single coral colony is comprised of 100s to millions of individual “polyps” (seen above). Each polyp can be visualized as an upside down jellyfish (coral’s close cousins) with their backs cemented to a surface and tentacles extended outward to capture passing food particles, live prey, or new symbionts. However because coral live in nutrient depleted environments, in addition to filter feeding, polyps harbor single-celled photosynthesizing symbionts inside their cells. Those symbionts (aka zooxanthellae) typically provide ~90% of the coral’s energy needs. Just 40 years ago it was believed all corals were host to just one photosynthesizing symbiont, a single species from the dinoflagellate genus Symbiodinium. But thanks to technological advances in genetic sequencing, we now know a coral species can harbor several potential species or types of Symbiodinium algae, each capable of responding optimally to a different set of environmental conditions and coral physiology. As predicted by the adaptive bleaching hypothesis, improved genetic techniques have revealed a wondrously diverse community of symbionts that coral can choose from. Coral can no longer be viewed as organisms that only adapt slowly over evolutionary millennia via genetic mutation and natural selection. Coral must be seen as an “eco-species” (aka holobiont) that emerges from the synergy of the coral and its varied symbionts. And we now know those emergent eco-species can rapidly evolve with changing climates by shuffling and shifting those symbionts.
A single colony’s polyps are typically all clones resulting from asexual reproduction and on their own offer the colony scant genetic versatility. However within a colony, a wide variety of symbionts can be harbored within a small percentage of polyps, although one symbiont type typically dominates. That small percentage of “cryptic” polyps often survive severe bleaching episodes and then multiply rapidly over the skeletal remains in a process known as the Phoenix effect. Just one square centimeter of coral tissue typically harbors a million individual symbionts and on average those symbionts can double every 7 days. Thus after severe colony bleaching, a more resilient colony can arise in just a few years with better-adapted symbionts now dominating. Likewise symbiont variability within a reef results in some colonies bleaching while adjacent colonies of the same species do not. And similarly a varied symbiont and coral community allows neighboring reefs to adapt to their unique regional climates.

Figure 6 Colony on the left remains unbleached
Variations in coral reproduction can conserve an “ecospecies” or rapidly promote greater ecospecies diversity. Twenty-five percent of the coral species produce larvae inoculated directly from their parent’s symbionts. However 75% of the species produce larvae that initially lack a symbiont. Only after coral larvae settle on a surface, do those larvae engulf one or more different types of free-living Symbiodinium, drawing them inside their cells. As the larvae develop into mature polyps, coral typically keep the symbiont types best suited to the local microclimate and expel the others. In this manner completely new eco-species emerge.
Furthermore as conditions change, all species can shuffle their symbionts as polyps will expel their current residents and acquire a different type that had been harbored by a neighboring polyp. A colony can also shift its symbiont population by acquiring new types not yet hosted by the colony but are present in the reef. Due to improving genetic techniques, previously undetected types of symbionts with greater thermal tolerance are now being detected after bleaching events. Thus a combination of symbiont shuffling and shifting is the key to corals’ rapid adaptation. Although bleaching can result in coral death due to starvation when new symbionts are not acquired quickly enough, surviving polyps with their altered symbiont community have the potential to re-direct the reef on a trajectory that is better suited to the new environment. Or if conditions return to those prior to an extreme event, coral can re-acquire their old symbiont types.
Scientists have found that coral colonies nearer the surface often harbor a different type of symbiont than colonies living just a few meters deeper. The symbionts residing closer to the surface may be better adapted to high irradiance by making proteins that protect against too much ultra violet light or by modifying their photosystem. Conversely symbionts living at greater depths may photosynthesize more efficiently under low light conditions but are more susceptible to UV damage. Transplant experiments revealed that when coral colonies growing at greater depths were relocated closer to the surface, the polyps expelled their symbionts resulting in temporary bleaching. Bleaching allowed polyps to acquire new symbionts better adapted to higher irradiance. However colonies adapted to high-light surface conditions, photosynthesized much more slowly when transplanted to lower depths. Bleaching never happened and the coral died. Although experiments can force bleaching by raising temperatures, other controlled laboratory experiments found that in the absence of stress from high solar irradiance, anomalous temperatures 4 degrees above average still did not induce bleaching.
According to the adaptive bleaching hypothesis we can infer that bleaching events are not simply the result of recent global warming. Bleaching should have been ongoing for millions of years, as background temperatures have risen and fell. Thus we would expect that as the Little Ice Age ended and naturally temperatures rose, there should be observations of bleaching in the early 1900s. And indeed there are albeit limited. For example bleaching was reported in Florida on hot days in the early 1900s. But more telling, enough warm weather bleaching had been observed in the early 20th century that the Great Barrier Reef expedition of 1928-29 focused on warm weather coral bleaching when oceans were cooler than today and long before any possible CO2 warming effect.
Coral Response to Climate Change
Since his first Greenpeace-funded 1999 study, Hoegh-Guldberg has promoted catastrophic climate change as the biggest threat to coral reefs. His papers are frequently cited as evidence of climate related coral demise by some researchers and hyped by media outlets that boost readership by promoting climate catastrophes. The bases for his claims relied on 3 simplistic assumptions that a) bleaching is evidence that coral have reached their limit of maximum thermal tolerance, b) bleaching will increase due to global warming, and c) coral cannot adapt quickly enough to temperatures projected by climate models.
In 1999 Hoegh-Guldberg argued “thermal tolerances of reef-building corals will be exceeded within the next few decades” and coral reefs “could be eliminated from most areas by 2100” due to climate change. In his 2014 paper he continued to dismiss the emerging science supporting the adaptive bleaching hypothesis, belittling it as a “persistent mirage”. His catastrophic claims also intensified, suggesting “as much as 95% [of the world’s coral] may be in danger of being lost by mid-century.” To support his extirpation claim he cited two of his own previously published papers. Hoegh-Guldberg’s history of exaggeration and circular reasoning has led other coral experts to accuse him of “popularizing worst case scenarios”, while others have accused him of persistently misunderstanding and misrepresenting the adaptive bleaching hypothesis. Furthermore other researchers have pointed out the pitfalls and weaknesses in framing threats to coral based on a simplistic temperature threshold. They argue, “A view of coral reef ecosystems that emphasizes regional and historical variability and acclimation/adaptation to various environments is likely to be more accurate than one that sees them as characterized by stable and benign temperature regimes close to their upper thresholds.”
Improved genetic sequencing is increasingly providing evidence that in response to warm water bleaching events coral begin acquiring new heat resistant symbionts. The results below from Boulotte 2016 show that over the course of 2 years, colonies radically altered their symbionts. The pie charts represent the changing percentage of dominant symbiont types due to shuffling in a single reef species. The bar graphs list just the rarer symbionts and stars identify types not previously detected suggesting an ongoing shift. Symbionts “types” are characterized first by their genetic lineages known as clades. When the adaptive bleaching hypothesis was first proposed, only 4 clades were known. Now at least nine have been identified. The most heat resistant symbionts belong to clade D, but other heat resistant types have evolved within other clades. Many earlier acclimation studies simply identified a symbiont’s clade. But we now know each clade can harbor hundreds of types (potential species) and improved detection of those species is uncovering more shifting. The most heat resistant species identified to date belonged to clade C. As seen here, different types/species are identified as D_I:6 or D1.12. As illustrated below after 2 bleaching episodes, a new symbiont species from clade C began to dominate and previously undetected clade D symbionts began to appear more frequently in just 2 years.

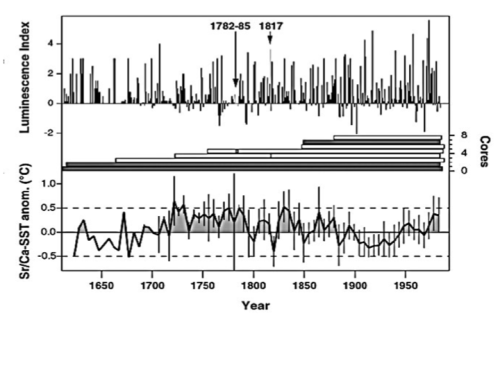
Nevertheless Hoegh-Guldberg 2014 continues to dismiss coral’s ability to rapidly adapt arguing, “current rates of change are unprecedented in the past 65 Ma [million years] if not 300 Ma.” But such exaggeration is pure nonsense. Ocean temperatures were warmer just 1000 years ago, and paleo-studies of temperatures in the Great Barrier Reef suggest local reef temperatures were higher between 1720 and 1820 as illustrated below from Hendy 2003. (Their luminescence index measures changes in salinity associated with monsoons). Perhaps CO2 concentrations are higher now than over the last 300 Ma. But given the extreme warmth just 65 million years ago, that is evidence that our climate is not very sensitive to CO2 concentrations, as realized by more researchers. In contrast to IPCC models that predict more warming that Hoegh-Guldberg ties to coral demise, climate experts note the Holocene temperature conundrum. While CO2 driven models simulate 6000 years of warming due to rising CO2, all the proxies indicate a cooling trend interrupted only by warming spikes.
Although coral genomes may evolve slowly, their symbionts have extremely fast generation times, averaging every 7 days. Furthermore the symbiont community consists of hundreds of symbionts that have already adapted to a wide variety of temperature, irradiance and salinity variables within different microclimates over the past million years. Symbiont shuffling and shifting is an evolutionary masterpiece that circumvents plodding evolutionary mechanisms of most organisms with long generation times and enables immediate adaptation.
A good summary statement is provided by Baker et al. “flexibility in coral–algal symbiosis is likely to be a principal factor underlying the evolutionary success of these organisms”.
Biosketch: Jim Steele is Director emeritus Sierra Nevada Field Campus, San Francisco State University and author of Landscapes & Cycles: An Environmentalist’s Journey to Climate Skepticism
JC note: This post was submitted via email, I did some minor editing on it. As with all guest posts, please keep your comments civil and relevant.

{kind=link}
Pingback: Coral bleaching debate – Enjeux énergies et environnement
A magisterial denouement of the coral bleaching dystopia-de-jour. Just as good at the second viewing (after WUWT). Thanks again Jim! As for the bleaching doomsayers – you’ve exposed their dirty laundry, and hung them out to dry!
I was curious how the skeptics could deny the obviously damaging impact of anthropogenic climate change on the Great Barrier Reef. Amazing number of contortions and distortions littered throughout this propaganda piece.
“There is a perception that bleaching suddenly became more common only since the 1980s, leading some to speculate bleaching is due to rising CO2 and global warming.”??? There is not a perception, there is reality that bleaching is more common since the 1980’s. Some speculate? I guess that is true if “some” means the overwhelming consensus of peer-reviewed science. In 1987, marine biologists had never even seen or heard about regional coral bleaching: http://www.nytimes.com/1987/12/15/science/experts-are-puzzled-by-widespread-coral-bleaching-in-caribbean.html
The peer-reviewed science is overwhelming and clear that coral bleaching due to elevated ocean temperatures is happening more often and is projected to get worse.
Referring to other times in earth’s history when the climate was hotter or colder is a favorite meme of climate denialists. Of course, pointing out that fact ignores that man is now changing the climate and that the rate of change is too fast for natural adaption to catch up. Current climatic warming is occurring much more rapidly than past warming events. When global warming has happened at various times in the past two million years, it has taken the planet about 5,000 years to warm 5 degrees. The predicted rate of warming for the next century is at least 20 times faster. http://earthobservatory.nasa.gov/Features/GlobalWarming/page3.php
While marine biologists are hopeful that some corals may be able to shuffle symbionts and withstand hotter oceans better, it is folly and pure manure to imply that this “enables immediate adaptation.”
This article cleverly distorts many issues to try and pretend that coral bleaching is not a problem, but like most climate denier propaganda, it is counter-productive and wrong. The fact that 35% of the Great Barrier Reef is dead or dying due to hot water is just a fact that must irritate this those that think that AGW is a hoax, and/or is not a problem. Corals are dying due to climate change and distorting science, as done by Mr. Steele, does not change that.
if 100% of that 35% is due to CO2 and it will take 100,000 years for current CO2 levels to drop to normal, saving the GBR looks a bit like a lost cause. Might as well pick another battle.
Now if some of that 35% is due to something other than CO2, perhaps that might be worth looking at. Why there is 65% not dead might be worth looking at. But is it is all due to CO2, kinda of a waste of time protecting the GBR from anything else is there
Lots of bleaching, lots of healthy coral. Severe in stretches of the north, not bad at all in many tourist areas, where, as I heard from one eyewitness, the bleaching was very severe in the late 1970s. Of course, the ’98 event was rough.
An insight into the activist mind:
http://www.2gb.com/audioplayer/172821
Don’t show them anything healthy or pretty. They want to go straight to the leper ward to dispense their concern.
Tripp Funderburk: “Amazing number of contortions and distortions littered throughout this propaganda piece.”
http://bitsocialmedia.com/wp-content/uploads/2013/07/Internet-Troll.jpg
Here is data from a 1997 paper showing 1) bleaching events before 1970 are unknown due to lack of observations and 2) observed temperatures were higher in the 1930s. However this paper was published in 1997 and missing the most recent temperatures which may have matched that period. But due to the hiatus recent temperatures have not likely exceeded that earlier time period
http://landscapesandcycles.net/image/115712066.jpg
http://www.int-res.com/articles/meps/158/m158p289.pdf
Much of the bleaching hysteria is the assumption that there is more bleaching now, but due to observational bias that assumption is mere speculation
Tripp,
I extend the same $500 bet to you. While Hoegh-Guldberg suggests by mid century we will lose 95% of our coral by 2050, I will bet you that in uninhabited islands where reefs are not assaulted by over fishing and landscape changes and thus primarily climate change, that those reefs (Smith 2016) will suffer no more than 5% mortality and will show signs of recovery
This post from Jim epitomizes how he distorts the conclusions from the studies that he cites.
Jim Steele | May 31, 2016 at 3:36 pm |
Here is data from a 1997 paper showing 1) bleaching events before 1970 are unknown due to lack of observations and 2) observed temperatures were higher in the 1930s. However this paper was published in 1997 and missing the most recent temperatures which may have matched that period. But due to the hiatus recent temperatures have not likely exceeded that earlier time period
http://www.int-res.com/articles/meps/158/m158p289.pdf
Much of the bleaching hysteria is the assumption that there is more bleaching now, but due to observational bias that assumption is mere speculation
<<<<<<<
So, Jim cites this paper and his take-away is that observed temperatures were higher in the 1930's. However, the paper clearly states in the initial abstract that:
"There has been a significant increase in annual summer and winter air temperatures in the Magnetic Island area since the middle of the present
century. Significant warming trends have also been observed in the nearby state of Queensland and in Eastern Australia over the same period."
The paper also states:
"It has been established that corals in subtropical and tropical locations are existing very close (1 to 2°C) to their upper thermal lethal limit during
summer months (Coles et al. 1976)"
and
"The relationship between bleaching events and periods of elevated air and seawater temperatures (Figs. 1 & 2), and the absence of similar air temperatures from the late 1930s until the late 1980s, is suggestive that
bleaching events at Magnetic Island are a comparatively recent phenonenon (in the last 25 yr) and are associated with a change in the local climate system."
Jim claims that the paper says that observed temperatures were higher in the 1930's when the entire thrust of the study is that there had been significant warming trends and that bleaching is a recent phenomenon due to rising temperatures. Jim pulls out one graph from one location that has one or two outlier high temperatures in the 1930's to distort the entire thesis of the paper. This is just one of dozens of examples of Jim concocting his own conclusions that are in direct opposition to the conclusions to the studies that he cites.
Tripp, You keep revealing that you can not read carefully and are consumed with sniping.
NIce try there is no argument that there has been a warming trend since 1960. You try to accues of of distorting the paper so lets look at their graph again. Look carefully and notice the high temperatures in the 1930s. That warmth was a common phenomenon around the world.
There is absolutely no conflict between what the authors and I have said. That paper did say
So Tripp turn your attention to the graph and notice that after the peak in the early and mid 30s, temperatures dropped. We would all agree that if the coral symbioses were at their thermal limit in the early 1930s, then a drop in temperatures would ease that stress. I do not disagree that recent warming once again brought the symbioses to that thermal limit.
What I am arguing is that if bleaching is a function of reaching their thermal limit than they must have bleached in the early 1930s as well. That bleaching was not observed is a function of no one looking, as the authors admit. If so then recent bleaching is not something new forced by CO2, but just past of the natural oscillation processes. But whether or not there was bleaching is now all speculation and we can only infer what must have happened.
If Magnetic Island coral actually did not bleach in the early 1930s despite similarly high temps, then there must be another reason for recent bleaching because similar temperatures did not cause bleachig. That’s the conundrum.
More personal attacks simply show you are incapable of critically analyzing the conundrum. But I suspect so and that’s why I named it Tripp’s Temperature Conundrum
https://i2.wp.com/landscapesandcycles.net/image/115712066.jpg
Thank you Thunderbunk for trotting out the party line just in case we’d forgotten.
Tripp, You do a lot of “denier” name calling and you appeal to a consensus but you offer very little scientific evidence or substantive arguments.
You link to a NY TIme article from 1987 where scientists were perplexed by bleaching, as if that refutes by argument that bleaching is as been quite common in the past but there was not n adequate global observational system. The 1980s Caribbean bleaching came on the heels of widespread damage from white band disease and an El Nino event.
Indeed there is widespread consensus that warm El Nino events have triggered widespread bleaching, but there is little evidence that CO2 has any effect on El Nino frequency or intensity. The climate models are all over the place, and those who wish to implicate CO2 cherry pick only the models that suggest CO2 will make El Ninos worse. But according to the IPCC’s science basics the tropics are least sensitive to a CO2 greenhouse effect due to the high humidity and convection. And NASA data for the Nino 3.4 regions shows no warming trend as seen in their graph here
http://data.giss.nasa.gov/gistemp/2011/Fig8.gif
The lack of a temperature trend in NIno waters suggests that other factors have made coral more susceptible to the same natural El Nino events. Furthermore as the essay also illustrates, El Ninos were more common over the past 3000 years. To leap from the undeniable observations that bleaching has been associated with warm waters from El Nino and then argue its all due to rising CO2 climate change is a huge leap of logic based on unsupported speculation. As Smith 2016 has shown, coral surrounding uninhabited islands are doing very well and look as good as they did 1000 years ago. If global climate change was the driver of coral mortality, we should expect both inhabited and uninhabited islands to suffer similarly.
Perhaps you can explain why scientists were also perplexed much earlier by warm weather bleaching and launched the Great Barrier Reef expedition of 1928-29 when oceans were cooler than today and long before any possible CO2 warming effect.
You then continued with more insults such as”Referring to other times in earth’s history when the climate was hotter or colder is a favorite meme of climate denialists. ” But I suppose you did not take many science classes en route to your MBA. It is common practice in biological and ecological research to frame a problem within its evolutionary context, but perhaps you blindly think all biologists who do so are “deniers”.
If coral survived 1C warmer oceans just 1000 years ago and 2.1C warmer waters 6000 years ago, we can reach one or two reasonable conclusions. 1) coral have mechanisms to adapt to warmer temperatures than today and should be able to do so in the future. OR 2) coral have been evolving in such a way to survive colder oceans of the Little ICe Age and the slightly warmer but relatively cooler Holocene temperatures of the 21st century. If you believe in #2 and the traditional slow evolutionary processes it suggests corals’ genetic capability of adapting to warmer water of the past has been culled by the LIA but it then becomes very difficult to understand how coral have survived all the past variabillity between glacials and interglacials. Perhaps you can explain how CO2 warming has caused coral to also bleach during cold snaps and in places with cold upwelling?
Rapid symbiont shuffling and shifting explains how coral can be adapted to a very narrow temperature ranges yet still survive frequent variability. To dismiss an analysis that includes an understanding of how coral have evolved and call it the work of deniers, suggests you are not a scientist and perhaps your criticisms are due to politics or perhaps your business thrives due to climate fears so your financial blinkers prevent you from seeing the bigger picture.
I linked a story that described that the best marine biologists had never seen regional coral bleaching and did not even know what was happening in 1987. Now bleaching happens nearly every year. It is happening more often because global temperatures have risen due to CO2 emissions. You write: “little evidence that CO2 has any effect on El Nino frequency or intensity.” That is just ridiculous. Do you deny that average global temperatures have risen about 1 degree celsius since 1880? Is it not entirely obvious that if the baseline temperature has increased, then El Nino events will be more severe and hotter and more damaging to corals? Do you imply that the Great Barrier Reef expedition was launched to study coral bleaching? Do you have some citations for this?
You also ignore that bleaching makes corals more susceptible to disease and likely have played a role in the White Band disease decimating Caribbean corals.
Also, the rapid pace of temperature increase due to man-made greenhouse gas emissions (do you deny this reality?) is many times faster than in the past. There are about 8 degrees celsius between glacials and interglacials over 100’s of thousands of years. We just experienced 1 degree in 140 years due to man’s greenhouse gas emissions. Corals can’t adapt to that rapid change, despite your preposterous claim of “immediate adaptations” miraculously provided by symbiont shuffling.
Shifting symbionts is an interesting area for study and hopefully will help corals recover from massive global bleaching and death caused by man-made global warming. However, your post seems more focused on pretending that global warming is not a problem than a serious attempt to understand how symbiont shuffling could help corals survive this massive bleaching and die-off. That is just the truth.
Jim, I’ve been noticing (not always reading) death-of-GBR stories since they were a good filler for Sunday newspapers in the 1970s.
Rest assured, as coral cover increases again, the subject will just be choked back to simmer till the next bleaching event.
Conservationist Ben Cropp is Australia’s best known diver and at eighty still dives regularly about the reef. He says reef damage of all sorts is real but consistently beaten up out of all proportion.
Australia’s largest oceanic reef system, Scott Reef off WA, was hit really hard in ’98, with 80% bleaching. Full recovery to pre-bleaching levels took about twelve years.
Don’t know why we fuss, since bleaching is probably meant to happen, like leaf fall and bushfire. It’s odd that the people most vocal about climate change don’t actually believe in climate change. Like a reef is just going to sit out there in ravaging nature and stay static and pretty for them! No wonder they swallowed the hockey stick.
This is just gibberish: “The climate models are all over the place, and those who wish to implicate CO2 cherry pick only the models that suggest CO2 will make El Ninos worse.”
All the models, all the evidence, all the thermometers indicate temperatures have risen and will continue to rise due to CO2 emissions. Rising baseline temperatures will make El Ninos hotter and more damaging to corals. How on earth is that cherry-picking? This is pretty basic stuff about the greenhouse effect that you seem to be denying?:
-If ppm of CO2 continues to rise, the earth will get hotter.
-If it gets hotter overall, then El Ninos will be even hotter and more damaging to corals.
-35% of GBR just died due mainly to hot water.
-2016 has seen record-breaking heat that caused global coral bleaching and widespread mortality.
-The widespread death of coral reefs and the ecosystems they provide is a bad thing.
Do you dispute any of that?
Your agenda and tactics are clearly displayed in this description of the consensus surrounding climate change: “those who wish to implicate CO2.” “Those” includes National Academy of Science, NASA, NOAA, DoD, 175 nations that signed Paris Accords, the Pope, and 97% of climate scientists. Boy, there sure are a lot of cherry-pickers. I have read your climate denial diatribes on websites across the internet. Your posts are repetitive, unpersuasive, argumentative and are easily refuted time after time.
It is a shame that you devote your time and energy to try and diminish the obvious damaging impacts of AGW on coral reefs. What we are seeing is neither “normal” nor “temporary”. Identifying one adaptive strategy that some corals might posses does not lessen or refute the catastrophic loss of corals due to AGW. And skeptic propaganda to diminish this tragedy does not help us find solutions to reduce the greenhouse gas emissions and future temperature increases that will kill more coral.
Tripp
As regards the likely cause and effect of coral beaching, it is surely worth trying to look back further in time than the 1998 event?
http://www.int-res.com/articles/meps/158/m158p289.pdf
See figure 2 for sporadic temp data for Magnetic Island coral reefs back to the 1930’s. The period generally from around 1918 to around 1940 were very warm globally then dropped back in the ‘global cooling’ scare that lasted until around the early 1970’s. This seems to be mirrored in the graphic.
I found this article interesting- but appreciate the bearer of news might not have the credibility you believe warmist scientists might have. I found it intriguing-but can’t confirm-that these corals suffering bleaching also grow in Papau new guinea where the waters are 2 degrees C warmer
http://jennifermarohasy.com/2012/07/activist-scientists-crying-wolf-on-coral-bleaching-and-climate-change/
The comments in the article are educational. This one resonated with me, because as a historical climatologist, I often find that large climate edifices are built on the flimsiest of foundations
“The facts: Nobody knew a thing about the GBR until after WW2 and the Battle of the Coral Sea, after which SCUBA aqualungs became available outside military use and people began to explore coral reefs in any close detail, notably Jacques Cousteau.. How can so-called experts forecast the future of coral reefs from such fledgling research?’
The words ‘we don’t know’ rarely seem to appear in any scientific papers. The past –prior to satellites-is often a closed book to many researchers.
We know very little of the past performance of reefs in the warm times that existed prior to the 1990’s. I chronicle many of these and mosomoso often does this in respect to Australia.
tonyb
Tripp says
Tripp attempts to denigrate again but with little evidence,
Tripp you seem to get all your claims from media hype and ignore the science. All those squiggly lines that look like a 2nd graders scribble are climate models predictions. Note how they are all over the place and wose disagree with satellie and radiosonde data
http://www.drroyspencer.com/wp-content/uploads/CMIP5-73-models-vs-obs-20N-20S-MT-5-yr-means1.png
And you seem to be denying the data showing no trend in temperatures in the NIno region, so I will post the graph again
https://i2.wp.com/data.giss.nasa.gov/gistemp/2011/Fig8.gif
You write: “little evidence that CO2 has any effect on El Nino frequency or intensity.” That is just ridiculous.
Tripp doestn know El Nino. Scientists say
Climate models that are forced with increased greenhouse gas concentrations simulate changes in ENSO variability but importantly, the models do not agree on the sign of change nor on the mean state of the tropical Pacific Ocean
http://onlinelibrary.wiley.com/doi/10.1029/2011PA002139/abstract
>>>>>>
Jim, I know you rely on semantics and splitting hairs to support your discredited climate denial posts. However, it is just obvious that if global average temperatures have increased, then additional increases caused by El Nino will be even worse than the El Nino’s in the past. Either you are dumb, or purposefully obtuse if you do not see this obvious issue of overall average temp increases making El Nino events even more damaging to corals.
Tripp asks, “Do you deny that average global temperatures have risen about 1 degree celsius since 1880? ”
Of course i believe temperatures rose since the LIttle Ice Age. But most climate scientists suggest that CO2 did not reach concentrations capable of causing significant changes in temperature until 1950.
Does Tripp deny there were 100 years of natural causes for that temperature increase. Does Tripp those natural causes stopped in 1950? Is Tripp aware that there is an unsettled debate about our climate’s sensitivity to rising CO2?
>>>>>>
Jim, there may be some debate over “sensitivity” and the exactness of computer models trying to predict variability, but there is no debate by the vast, vast majority of legitimate scientists about man-made emissions causing temperatures to rise:
Statement on climate change from 18 scientific associations:”Observations throughout the world make it clear that climate change is occurring, and rigorous scientific research demonstrates that the greenhouse gases emitted by human activities are the primary driver.” (2009) http://climate.nasa.gov/scientific-consensus/
You refer to the consensus as ” “those who wish to implicate CO2.” This means you don’t wish to implicate CO2 in climate change. This makes you a climate denier and irrelevant to legitimate science.
Jim, why do all your temperature graphs leave off the record-setting years in 2014, 2015, and 2016. Are you just dusting off your old denial posts, or are you purposefully hiding these record-setting years so you can cling to the hiatus myth?
Jim, your sad attempt to downplay the devastation to coral reefs is shameful and baldly transparent. You are agenda-driven and only seek to minimize an obvious and dangerous tragedy on-going on the GBR. You can’t have your denial essays published in legitimate science publications (where they would be rejected by legitimate scientists), so you engage in subterfuge and mud slinging on numerous message boards across the internet. Good luck with that.
Tripp you are losing all credibility as your replies to scientific are increasingly irrational and trollish. You are denying the evidence.
I report from peer reviewed science that
Tripp’s “cogent reply” denies the science and launches more name calling hoping to divert the issue
“Jim, I know you rely on semantics and splitting hairs to support your discredited climate denial posts. However, it is just obvious that if global average temperatures have increased, then additional increases caused by El Nino will be even worse than the El Nino’s in the past. Either you are dumb, or purposefully obtuse”
I ask
Tripp avoids the facts and blathers about consensus. Everytime Tripp wild claims are refuted, he steps up the personal attacks and name calling.
Ask Tripp how coral survived the more rapid temperature changes of past, and all he can say is you are a denier.
Tripp your twisted meme and train of logic is:
1.CO2 is rising and is a greenhouse gas, (we all agree that)
2. Therefore oceans must warm beyond corals’ limits (history shows you are wrong at least for the next 2.1C rise)
3. Therefore discussing coral’s adaptation mechanisms are just a denier’s ploy. (so Tripp diverts the focus away from their marvelous adaptation mechanism.
Tripp I would never hire your company to do coral restoration work, if you ignore all the relevant issues regards coral biology. What do you tell clients? Drive a Prius and the coral will be save? LOL Does the help restore the 95% of the reef mortality from cyclones nd predators and disease?
So tell me Tripp why are all those coral around uninhabited islands doing so much better?
Thunderbunk that explains why there are so many coral reefs in the arctic regions. Thank you also for demonstrating why a whole lot of pretty words strung together can overcome reality. Viva word-based arguments viva.
You state things that simply are not true. You state that, “Bleaching is often temporary and normal.” We have had 3 global bleaching events, the first starting in 1998. These events have coincided with El Ninos combining with the elevated average temperatures caused by man’s emissions of greenhouse gases. So, these events are not “normal” despite your assertions and desire to diminish such a clear and dramatic example of the damages of climate change. What’s more, increases in average global temperatures are expected to be within the range of 0.5°F to 8.6°F by 2100, with a likely increase of at least 2.7°F for all scenarios except the one representing the most aggressive mitigation of greenhouse gas emissions. (IPCC (2013). Climate Change 2013: The Physical Science Basis). So, some corals might adapt using symbiont shuffling or other strategies, but these corals cannot adapt to such rapidly rising temperatures and increasing frequency in bleaching events.
It is estimated that 19% of the world’s coral died in 1998. That is not temporary. 35% of the corals on the GBR are dead. That is not temporary. And you admit that lack of recovery is a concern. So, how can that be “temporary and normal?”
You seek to normalize global coral bleaching and death so that you can pretend that it is not what it is: dramatic and clear evidence of the damaging effects of man-caused climate change and global warming. Identifying one survival strategy does not mean rising temperatures.
The only part of your analysis that is relevant and true is your quote from Hoegh-Guldber: “a) bleaching is evidence that coral have reached their limit of maximum thermal tolerance, b) bleaching will increase due to global warming, and c) coral cannot adapt quickly enough to temperatures projected by climate models.” You did not refute any of that.
TF:
Could you provide citations for your numbers? For example, the likely source of your “35%” was the estimate from 84 reef site surveys along the GBR which includes “dead” and “dying” in its totals. It also found bleaching concentrated in particular areas. Is that the source of your blanket assertions?
And, to be clear, do you assert these cited bleachings = permanent destruction of these coral reefs? That also would seem to be an exaggeration.
https://www.coralcoe.org.au/media-releases/coral-death-toll-climbs-on-great-barrier-reef
Tripp you seem woefully unaware. It is absoutely true that “Bleaching is often temporary and normal.” Coral typically reduce symbiont density in the summer and increase them in the winter. Often that results in minor bleaching.
The 1998 event indeed result in greater bleaching and mortality but that does not refute normal and temporary bleaching.
Tripp you argue that global warming causes hotter water that causes more bleaching but the fact that there is no temperature trend for the El Nino 3.4 region suggests it is not any rise in El Nino water temperature that is the problem. Why does Tripp avoid discussing facts that contradict is fears?
https://i2.wp.com/data.giss.nasa.gov/gistemp/2011/Fig8.gif
Regards your empty assertions that I did not refute HOegh Guldberg’s 3 assumptions
Bleach is a sign coral have been stressed. Coral beach during cold snaps and clear sunny days when water temperatures have not reach the coral’s maximum. Thus the assumption of reaching maximum thermal tolerance is speculation that assumes a new symbiotic partnership will not transpire.
This is based the myopic view from the first assumption. All major bleaching events have been caused by El Nino events. and there is no evidence that El Ninos will get worse or more frequent, or that there was less bleaching in the recent past when El Nino events were far more frequent
More speculation that denies all the evidence that symbiont shhifting can result in the acquisition of heat tolerant symbionts over the course of just a few years. It also denies that coral were able to adapt during past rapid climate change, change far more rapid than in recent decades.
Tripp has erroneously assumed that rising temperatures since the end of the Little Ice Age have been driven by rising CO2, but other researchers have revealed an equally(greater) high rate of natural temperature rise in the 1920s and 30s, with peak temperatures greater than at the end of the 20th century, as shown from data for Magnetic Island.
Furthermore the bleaching status of those reefs was unknown due to lack of an observational system.
http://landscapesandcycles.net/image/115712066.jpg
1) If there were no bleaching events in the 1920s and 30s despite those higher temperatures, then temperature is an unlikely primary driver of bleaching.
or
2) If there was bleaching during those higher temperatures, then coral survival must have been due to rapid adaptation such as symbiont shuffling and shifting. The temperature change from the cold LIttle Ice Age to 1930 was at a far faster rate, than from 1930 to 1997 when there was no warming trend.
Jim,
The Magnetic Island data show only 4 warm years during the 20s and 30s relative to the 80s and 90s bleaching years.
In addition to the 5 bleaching events in the 80s and 90s, there were bleaching events in 1998, 2002 and this year – all El Nino years so likely warmer.
So the evidence is that frequency of warm events and bleaching events is increasing.
Steve
The magnetic island data starts only in 1927 so it would be reasonable to expect there was around another decade or so of warming prior to that.
However as the graphic points out, the bleaching status prior to 1970 is unknown. A unique bleaching hypothesis is being built on a shaky foundation
tonyb
Bleaching is correlated with warmth post 1970.
Our lack of evidence prior to 1970 does not weaken the evidence from this correlation given that the causation is also understood (even if bleaching can have other causes).
We know of El Nino events which were strong in effect for Qld – three or four in the first two decades alone of the 20th century, the ‘weak’ of 1902 being disastrous – and El Nino events which were strong globally (not just ’82-3 and ’97-8, but also the double whopper of ’40-2 when Pacific dwellers had their minds on other things than bleached corals. When the brute of 1914-15 came around there were similar distractions.)
Now, I agree that the marine biologist numbers were low and that aerial surveys were as rare as rocking horse poo in 1902. However, I doubt that reef damage was absent because biologists were absent from Australian reefs and the Wright Brothers’ first flight was a year in the future.
If a tree falls in the forest and nobody sees or hears it…it still falls, grasshopper.
By all means dismiss history and anecdote, but don’t then say things are “hotter”, “worse”, “more frequent” etc when you yourself have disallowed essential points of comparison.
Steve MIleswothy says
WRONG. The authors state that the bleaching status was unknown. ie there were no observations made, not that there was no bleaching. With those warmer 30s temperatures why wouldn’t it by more likely there was similar bleaching events
Steve MIlesworthy says
That’s why people say there are lies, damn lies and statisitics.
You should read the McClanahan paper about Indian Ocean coral, “Regional coral responses to climate disturbances and warming is predicted by multivariate stress model and not temperature threshold metrics”
“the relationship between warmer ocean temperatures and “bleaching has been equivocal and sometimes negative when the coolest regions were not in the analyses.” In other words coral living in the warmest waters were well acclimated to the warmest waters redistributed by an El Nino.
The evidence is that warmth-induced bleaching is increasing because the frequency of warm years is increasing and the impacts of the warmth are being sufficiently closely observed. That’s true even if the 5 warm years in the early 19th century caused unobserved bleaching events.
It’s interesting that your first “expert” whom you favourably quote also writes:
“Bleaching disturbances are likely to become a chronic stress in many reef areas in the coming decades, and coral communities, if they cannot recover quickly enough, are likely to be reduced to their most hardy or adaptable constituents.”
Steve
you commented about ‘ the five warm years in the early 20th century’ but these were only from 1927. Warming had commenced long before that elsewhere and it is very likely there was bleaching in this period but there was no one around to record it.
We can see evidence of very hot years throughout Australia’s brief history. The famous diaries from Watkins Tench in the 1790’s from the Sydney area demonstrated considerable warmth that caused parrots and bats to drop from the sky, a phenomenon claimed as unique and due to AGW when it happened just a few years ago
tonyb
http://onlinelibrary.wiley.com/doi/10.1111/j.1461-0248.2012.01861.x/abstract?deniedAccessCustomisedMessage=&userIsAuthenticated=false
Well Mosh, I can either merely rent the article for 6 dollars or buy the PDF for only 38 dollars. Which do you recommend I do?
Tonyb
https://sci-hub.bz/10.1111/j.1461-0248.2012.01861.x
AK
Your link led to a Russian site. Were you being satirical?
Tonyb
@climatereason…
No. It’s Sci-hub. AFAIK it will ask you to solve a capta (prove you’re a human) then get you the paper.
The link worked for me.
AK
Its entirely in Russian characters except for one green label which reads ‘Support the project’
What am I supposed to do?
Tonyb
I’m sorry, Tonyb, I can’t duplicate the problem from here, but if it’s the page I would expect you to see, there should be an entry field, with a picture of some letters above it. You know, the things that you use to prove you’re not a bot. If that’s how it is, just type in the letters you see.
If it’s something else, I’d say just go directly to https://sci-hub.bz and enter the doi in the entry field. It may not work in England, I get the impression some ISP’s are fiddling with sci-hub’s connections. If you still can’t get it to work, I’ll upload it to dropbox, but I don’t want to if I can avoid it.
tonyb:
The full paper “Evaluating life-history strategies of reef corals from species traits” by Darling, et al., is also available on researchgate.net.
AK and Opluso
Thanks for your help. No, there are no numbers or English letters anywhere on the site. It is just Russian! How curious. Will try one of the other methods suggested to locate it but is the article worth it?
Tonyb
Tonyb:
It may be more useful in a clubhouse debate about the best method for predicting species recovery and evolution under stress.
https://www.dropbox.com/s/m85jja5qwpixu28/darling2012.pdf?dl=0
too funny
“2) If there was bleaching during those higher temperatures, then coral survival must have been due to rapid adaptation such as symbiont shuffling and shifting. The temperature change from the cold LIttle Ice Age to 1930 was at a far faster rate, than from 1930 to 1997 when there was no warming trend.”
1. We have no good SST data before 1950. Just ask tonyb.. /sarc
and her comes the skeptic ASSERTING with no evidence
that temperature change from the LIA ( when and where) was
greater and had a faster rate..
2. There is basically no data on bleaching during that period or data
on what species were involved or how, if at all, they adapted, or perhaps were replaced with species more tolerant.
This is known in the business as making sh*t up.
@tonyb…
Hope the dropbox worked. Could you let me know? Thanks.
@Steven Mosher…
A novel paleo-bleaching proxy using boron isotopes and high-resolution laser ablation to reconstruct coral bleaching events Very preliminary so far…
AK
It doesn’t work on my iPad. I will try on my laptop in an hour or so and let you know. Thanks for your help.
Tonyb
http://phys.org/news/2015-02-la-nina-like-conditions-year-long-shutdown.html
La Nina-like conditions associated with 2,500-year-long shutdown of coral reef growth
If this is the same study I read about in a news story, they used PVC pipe to core sample coral reefs.
@AK
Very preliminary so far…
yup..
buut that doesnt stop skeptics from making stuff up when they need to
Makes me wonder if there are still the remains of coral along the shores of Lake Vostok, from about 450K years ago?
Apparently, exceptions are made for the paleo studies informing international policymakers. Good Enough For Government “Work”™
too funny™
AK
That dropbox link worked fine on my laptop. Thanks for your efforts
tonyb
Hope you enjoy it Tonyb.
Mosher says,
Are you saying the Magnetic Island data is unreliable??? Or you makin up sh*t that I extrapolated the warming trend to the whole ocean???
http://landscapesandcycles.net/image/115712066.jpg
Please explain why the rate of warming “at Magnetic Island from 1920 to 1930s, is NOT faster than the rate between the 30s and 1990s?
Jim Steele,
“Regional coral responses to climate disturbances and warming is predicted by multivariate stress model and not temperature threshold metrics”
Yes, but one of the stresses in the stress index is temperature, and the MSM model seems to have been fitted to the data by playing around with the parameters that measure the importance of each of its variables:
These 11 variables [in the Multivariate Stress Model] were synthesized using fuzzy logic algorithms, where functions that represent the effects these variables on coral bleaching response were estimated from field studies, applied as best-fit mathematical functions, and weighted based on the strength of the environmental-bleaching relationships
`“bleaching has been equivocal and sometimes negative when the coolest regions were not in the analyses.” In other words coral living in the warmest waters were well acclimated to the warmest waters redistributed by an El Nino.`
And those not in the warmest waters were not, and are at threat from warmer temperatures.
Yes it’s complex, but your case for dismissing the impacts of warming and climate change are based on very selective use of evidence.
The only dirty laundry exposed here is that Judith Curry provided a platform to spread disinformation. If Jim Steele is correct that shuffling symbionts provide miraculous “immediate adaptation” why is the Great Barrier Reef 35% dead, and remote, isolated reefs like Jarvis Island are 95% dead? If corals have this miraculous ability for immediate adaptation through symbiont shifting why are they all dead?
http://bigstory.ap.org/article/2534bee620964745ae90d2aa209ea356/scientists-vibrant-us-marine-reserve-now-coral-graveyard
“El Nino’s super warm water has turned what had been one of the world’s most lush and isolated tropical marine reserve into a coral graveyard, federal scientists said Wednesday.Researchers finishing an emergency undersea expedition found 95 percent of the coral dead around Jarvis Island in the Pacific Remote Island Marine National Monument . In November, much of the coral had bleached white but was alive.”
Jim Steele’s post is a pitiful attempt to cling to climate change denial. Remote, pristine reefs are dead due to hot water. Judith Curry posts Jim Steele’s spurious, out of context distortions to try and pretend that an increasingly hot planet due to greenhouse gas emissions is not a reality. I find it disgusting and shameful. Dead reefs due to record-setting temperatures due to AGW combining with El Nino are a clear example of the damaging impacts of climate change. This is not a game for deniers to selectively quote studies, reinvent history, and make ridiculous claims to try and pretend the obvious is not the obvious. Coral reefs are dying around the planet due to man’s Co2 emissions. Anything Jim Steele writes to attempt to obfuscate this reality is pure subterfuge and hogwash.
Tripp,
SST today at Laem Sing – 32.2 C.
Snorkelling over the local coral reefs continues. No current episodes of bleaching noticed. Maybe it’s not hot enough yet?
All wild populations increase and decrease. Many peer reviewed papers have come to the conclusion that population variations are chaotic, and unpredictable.
Most species that have ever existed on the face of the Earth now have populations of precisely zero – that is, they are extinct.
Good luck with saving the coral. Or the ammonite or the eohippus. Flora and fauna in the vicinity of hydrothermal vents can live in water temperatures of 113 C, and enormous pressure. No sunlight ever reaches them, of course!
If the seas start boiling, these flora and fauna will probably take over the oceans! No problem for Nature.
Worry all you want, but don’t expect me to pay. Are you interested in paying for things I consider important? I didn’t think so!
Cheers.
“Remote, pristine reefs are dead due to hot water.”
I can only find one variable here, temperature.
Wait another variable is CO2, and that makes temperature a result of CO2 and not a variable.
“The level of increased tolerance gained by the corals changing their dominant symbiont type to D (the most thermally resistant type known) is around 1–1.5 °C.”
“This is the first study to show that thermal acclimatization is causally related to symbiont type and provides new insight into the ecological advantage of corals harbouring mixed algal populations.”
http://rspb.royalsocietypublishing.org/content/273/1599/2305
Shifting symbionts apparently is being debated.
Tripp, so take my bet if you are sure of your analysis
I have seen nothing from you but sniping name calling, a sure sign you are incapable of arguing the evidence in a scientific manner.
Tripps critiques are the only pitiful posts. He says,
1. Regards Great Barrier Reef being 35% dead, on average on 5% of that mortality is attributable to bleaching which could be from warmth, from floods, from other stressors.
2. Different reefs bleach during different episodes for different reasons. And just because you labels Jarvis a a remote isolated island does not mean it has not suffered from human disturbances. In the 1800s it ws heavily mined for guano and during World War 2 the island was heavily bombed. It is now a reserve but that doesnt immediately alleviate disrupted landscapes. So what is your evidence that the only problem for Jarvis Island coral is rising CO2?
3. You persist in your catastrophic ranting via a snapshot in time. Again you must evaluate coral resilience and ability to adapt over periods of 2 or more decades. If a forest has a fire do you rant the forest is forever dead? Terrestrial vegetation is well adapted to fires and recovery to humans preferred state takes decades.
https://curryja.files.wordpress.com/2016/05/slide4.png
Thailand has moved to stop diving in many of its marine parks.
You have evidence that it is no longer the old radioactive waste from Bikini Atoll, or the boatloads of Cesium 137 etc…, dumping into the Pacific today? How much plutonium or left over MOX fuel does it take to kill coral reef? There must of been an NRC hearing on this subject and a paper trail of sorts should exits. Reading the paper is always more informative than just reading it in the newspaper. You must agree with me.
“Jim Steele’s post is a pitiful attempt to cling to climate change denial. Remote, pristine reefs are dead due to hot water.”
Utter, absolute, 24 carat, 100% scientifically illiterate rubbish.
You really haven’t a clue what you’re wittering about, and all the insults you can vomit up won’t change that.
http://bitsocialmedia.com/wp-content/uploads/2013/07/Internet-Troll.jpg
Jim Steele wrote, “Does Tripp deny there were 100 years of natural causes for that temperature increase. Does Tripp those natural causes stopped in 1950? Is Tripp aware that there is an unsettled debate about our climate’s sensitivity to rising CO2?”
Since the industrial revolution, man has been producing greenhouse gas emissions. “Data for the past 2000 years show that the atmospheric concentrations of CO2, CH4, and N2O – three important long-lived greenhouse gases – have increased substantially since about 1750. Rates of increase in levels of these gases are dramatic. CO2, for instance, never increased more than 30 ppm during any previous 1,000-year period in this record but has already risen by 30 ppm in the past two decades.”
http://www.acs.org/content/acs/en/climatescience/greenhousegases/industrialrevolution.html
Once again, Jim, you are full of it.
Here are some more facts:
Earth has warmed by 1.5°F (0.85°C)[1] Most of that warming has occurred since 1976. https://www.climate.gov/sites/default/files/Climate_gov_GlobalWarming_FAQ_15Aug13.pdf
Jim may want an unsettled debate, but it only exists on denier blogs and in his head. http://climate.nasa.gov/scientific-consensus/
“This is basically adaptation on speed.”
https://judithcurry.com/2016/05/24/coral-bleaching-debate/#comment-786298
And once again Tripp avoids answering with relevant facts. Due to the physics of CO2 most climate experts suggest CO2 had no significan ipact on global temeratures until 1950. Obviously Tripp denies physics in order to unleash his skeptic bashing.
And here is data showing a rapid rise of .8 C in ocean temperatures from 1900 to 1940
http://realclimatescience.com/wp-content/uploads/2016/04/Screenshot-2016-04-08-at-05.24.44-AM-down.png
Jim Steele wrote: “All major bleaching events have been caused by El Nino events. and there is no evidence that El Ninos will get worse or more frequent, or that there was less bleaching in the recent past when El Nino events were far more frequent.”
Here is direct evidence that refutes one of Jim’s many misstatements:
“Mass coral moralities in coral reef ecosystems have been reported in all major reef provinces since the 1870s. The frequency and scale of bleaching disturbances has increased dramatically since the late 70’s. This is possibly due to more observers and a greater interest in reporting in recent years. More than 60 coral reef bleaching events out of 105 mass coral moralities were reported between 1979-1990, compared with only three bleaching events among 63 mass coral moralities recorded during the preceding 103 years.
Prior to the 1980s, most mass coral moralities were related to non-thermal disturbances such as storms, aerial exposures during extreme low tides, and Acanthaster outbreaks. Coral bleaching accompanied some of the mortality events prior to the 1980s during periods of elevated sea water temperature, but these disturbances were geographically isolated and restricted to particular reefs zones. In contrast, many of the coral bleaching events observed in the 1980s occurred over large geographic regions and at all depths. ”
Jim also states, “bleaching events before 1970 are unknown due to lack of observations.” That is partially true, but there were 63 reports of coral mortalities before 1979 and only 3 were the result of bleaching. So, there were obviously some monitoring of coral mortalities and bleaching was not prevalent despite Jim’s misstatements.
http://www.marinebiology.org/coralbleaching.htm
Jim stated that
The link above is an excellent resource that accurately describes coral bleaching. It is fair, balanced and uses clear facts.
Tripp Funderburk: http://www.marinebiology.org/coralbleaching.htm
Here is a quote: Of the causing stressors of coral reef bleaching, many are related to local environmental degradation and reef overexploitation. Of the stressors mentioned above, only sea water temperature and solar irradiance have possible global factors driving changes and extremes. Global warming, along with ENSO events, change sea water temperatures. Ozone depletion increases the amount of UVR reaching the Earth’s surface, and possibly causing coral bleaching events.
Increased sea temperatures and solar radiation (especially UV radiation), either separately or in combination, have received consideration as plausible large-scale stressors. In most instances, wherever coral reef bleaching was reported, it occurred during the summer season or near the end of a protracted warming period.
You wrote: Jim Steele wrote: “All major bleaching events have been caused by El Nino events. and there is no evidence that El Ninos will get worse or more frequent, or that there was less bleaching in the recent past when El Nino events were far more frequent.”
Is it the phrase “All major” in the quote from Steele that you think is contradicted by the source that you quote “only sea water temperature and solar irradiance have possible global factors”?
Tripp Funderburk, thank you for the link.
Here is another quote: Most of the coral reef bleaching events of the 1980s occurred during years of large-scale ENSO activity.
Is it Steele’s “All” versus that source’s “most” that you are focusing on?
Tripp your sniping is getting desperate and more irrelevant. You say
“Here is direct evidence that refutes one of Jim’s many misstatements:”
http://www.theguardian.com/environment/2016/jun/07/the-great-barrier-reef-a-catastrophe-laid-bare?CMP=share_btn_tw How can the deniers deny the reality of the catastrophe caused by global warming and climate change considering the destruction of one of the earth’s most miraculous treasures? I really can’t find the words to accurately explain how despicable it is to pretend this is not a tragedy. Jim Steele, please think just a bit about how you have invested time and energy to pretend this is not what it is – a horrible tragedy caused by man’s carbon emissions. Judith Curry, please consider this horrific destruction when you allow charlatans to post nonsense on your blog. Do you have a semblance of honor or decency? Co2 is at record levels, coral reefs are dead because of it and the posters here make up pretend stories to cling to the myth of climate change denial. The reality of the death of one of the earth’s most precious ecosystems does not make you pause and consider that denying science and denying reality is somewhat misguided? I am sure the Jim and others will decry that this is only a news article and only obscure and distorted interpretations of legitimate science will be allowed. Bottom line, this blog and this post are attempting to pretend that the horrible reality of dying ecosystems is just a game and it is more important that deniers make up stuff to deny the obvious.
more nonsense :)
https://www.researchgate.net/publication/257685127_Coral_reefs_in_the_South_China_Sea_Their_response_to_and_records_on_past_environmental_changes
An estimated 80% of the corals in the south china sea have been destroyed in the past 30 years. Fairly regular bleaching events are document over the past 200 years and the recovery time was roughly 20 years until, “Australian Research Council Centre of Excellence for Coral Reef Studies, and shows the results of a survey of reefs off the coast of mainland China and in the South China Sea. The main driver the authors attributed to the loss was economic growth. That finding is supported by a recent WWF study that suggest China is using its environmental resources more than twice as fast as is sustainable.”
China building islands on coral reefs for military bases probably did help things.
“China building islands on coral reefs for military bases probably did help things.”
Irrelevant to the Thunderberks of this world.
You have to remember the axiom, “there are no enemies on the Left”.
Tripp,
First it appears that your emotional tirades and ugly personal attacks are likely the result of you being hoodwinked by exaggerated claims about reef destruction, along with your ignorance of rapid adaptation mechanisms via symbiont shifting as well as coral resiliency. Also I don’t think you read the whole essay nor the supporting citations. The essay is about emerging science. You seem consumed by an unsupported end of the world horror fantasy and a revealed lack of critical thinking skills about all the issues thoroughly discussed in the essay
I suggest you read the link posted by catweazle666
https://wattsupwiththat.com/2016/06/03/expert-scientists-exaggerated-coral-bleaching-story/
Corals survived millions of years of climate change. I just looked at the 1.5M year of the d18O Benthic Stack data (measure of global ice volume and deep ocean temperature) , may or may not be directly relevant, the most recent 0.8 Myr of data shows clear indisputable 100 Kyr Milankovic cycles
http://www.vukcevic.talktalk.net/d18O.gif
It is simpler than that. Tropical coral grows in less than 60 meters of water (it needs sunlight).
12,000 years ago the sea level was over 70 meters lower. 20,000 years ago it was about 120 meters lower.
All the current tropical corals are squatters who moved in since the end of the ice age.
Vuk
Very nice Milankovich signal, including the mid-Pleistocene revolution (MPR) where the alignment changes from obliquity to eccentricity – possibly representing a transition from strong to weak periodic forcing.
Tnx. Had to look up MPR
“The MPR is the term used to denote both the marked prolongation and intensification of these glacial-interglacial climate cycles that was initiated between 900 and 650 ka.
… After about 800 ka, glacialinterglacial cycles occur with a much longer mean period; approximately 100 kyr. Not only does the periodicity of glacial-interglacial cycles increase going through the MPR, but there is also an increase in the amplitude of global ice volume variations. The ice volume increase may in part be attributed to the prolonging of glacial periods and thus of ice accumulation.”
http://www.seao2.info/pubs/manuscript_maslin_and_ridgwell.pdf
Karner, et al. (2002) also detects the 100kyr cycle and has the added attraction of being co-authored by Richard Muller of BEST fame. However, their data does not show the recent decline seen in yours. That would seem to be an artifact of your moving filter?
http://muller.lbl.gov/papers/BenthicStack.pdf
Surely Great Barrier Reef’s short-form should be GBR, not GBF.
There are a couple of issues here:
1. The greens (who are liberal) should be encouraging this. A Great Barrier Reef goes against their whole philosophy. The fact that it is becoming an Average Inclusive Reef (AIR) should be treated by them as good news.
2. From the viewpoint of the more reality based, the real GBR problem is nutrient rich runoff which is providing food for the larva of the COTS (Commercial Off-The-Shelf) starfish. The starfish is what is killing the reef. They are even testing an robot starfish killer that injects them with vinegar.
Reefs (as I understand it) can only live in a nutrient poor desert since they can’t compete with starfish. If something isn’t done about the runoff or the starfish the reef will just have to move.
If periodic coral bleaching was the only thing man made global warming was responsible for then I’d just ignore it and write it off as the cost of having a better standard of living, But when the canary in the coal mine (coral) drops dead do we just hope we can find a more resilient breed of canaries or do we start worrying about the higher life forms that could be forced into extinction by the conditions that led to the death of the canary (coral) in the first place?
Scary, Jack. Did Halloween come and I didn’t get the memo?
When the canary in the coal mine invites 536 fellow canary species to come join him it doesn’t look like we are going to have canary shortage anytime soon.
Every time something, somewhere on Earth changes ( scary word ) , someone declares it to be a “canary” as if just saying the name of that bird proves something.
Life if a constant process of change. When things stop changing that will be the true canary.
jacksmith4tx: But when the canary in the coal mine (coral) drops dead
Why do you identify coral as the canary in the coal mine? Why not the more massive coccolithophore population, which is thriving?
We are lucky that coccolithophores have evolved to thrive in more polluted (phosphates) and acidic water. They may someday be one of our primary food sources when the fish are gone. Bon Appétit.
Things AGW causes:
The conditions that have led to increased bleaching appear to be the frequency of El Nino events that cause grave fluctuations between warm and cool temperatures. In contrast to the warm equatorial western Pacific with over 510 species, the cool eastern Pacific houses just 18 species.
Furthermore during the warm Holocene Optimum coral growth reached its max coinciding with reduced El Nino frequency, then began to decline as El Nino frequency increased over the last 3000 years. Today mass bleachings are all associated with El Ninos. Your coral canary species have not died, only individuals that are subjected to abrupt swings between warm and cold.
A magnificent post. The essence of the wonderful science core to CE.
Sadly, the ocean acidification/coral damage meme is underpinned by scientific misconduct by Fabricius et. al. concerning toxic hydrogen sulfide.
Essay Shell Games, but only in the ebook Blowing Smoke version. The guest post here of same name only covered oysters.
Great article!! Corals are in as much danger from “global warming” as polar bears.
Plus many. Had not thought of the analogy.
Lets put Jim Steele and Susan Crockford together editing an ‘endangement’ environmental anthology including their own writings. I would contribute essays No Bodies and Shell Games.
Jim Steele,
Thank you for a most excellent essay.
Cheers.
Jim Steele,
Let’s see what we can see …
Abstract
[1] Several negative feedback mechanisms have been proposed by others to explain the stability of maximum sea surface temperature (SST) in the western Pacific warm pool (WPWP). If these “ocean thermostat” mechanisms effectively suppress warming in the future, then coral reefs in this region should be less exposed to conditions that favor coral reef bleaching. In this study we look for regional differences in reef exposure and sensitivity to increasing SSTs by comparing reported coral reef bleaching events with observed and modeled SSTs of the last fifty years. Coral reefs within or near the WPWP have had fewer reported bleaching events relative to reefs in other regions. Analysis of SST data indicate that the warmest parts of the WPWP have warmed less than elsewhere in the tropical oceans, which supports the existence of thermostat mechanisms that act to depress warming beyond certain temperature thresholds.
Oops.
Yes, thanks. When most researchers start predicting an increased frequency of cold events, that will be especially salient information to consider.
Modeled estimates of global reef habitat and carbonate production since the last glacial maximum.
Get this pseudo-scientific mumbo-jumbo modelling crap out of here.
On a more serious note, more than just temperature affects rate of carbonate production. Here’s an interesting section:
2.6. Misperceptions
[…]
2.6.2. CO2 fertilization of zooxanthellae
A common misperception is that, similar to CO2-fertilization of land plants, increases in CO2 will enhance growth of the algal symbionts in corals, which in turn will lead to higher calcification rates. This misperception is based on two assumptions about the coral/algal symbiotic relationship: a) that increased photosynthesis increases calcification rates, and b) that zooxanthellar photosynthesis will increase with rising CO2. The first assumption is based on the fact that zooxanthellate corals calcify faster in the light than in the dark (estimated three times as fast by Gattuso et al. [1999]), and also that calcification in bleached corals slows or stops [Leder et al., 1991]. The inference is that photosynthesis enhances calcification. It should be noted, however, that the mechanisms for light-enhanced calcification have not been adequately resolved, and that the opposite – that calcification enhances photosynthesis – has also been suggested [McConnaughey and Whelan, 1997; Cohen and McConnaughey, 2003] (see Section 4.1). The importance of the second assumption is somewhat tempered by the fact that zoo-xanthellae primarily use HCO3– as a substrate for photosynthesis, and secondarily use CO2 (some of which is respired CO2). While aqueous CO2 concentrations will increase about 100% in concert with a doubling of atmospheric CO2, HCO3– concentrations will increase only about 14% (Figure 2). This increase in CO2 and HCO3– may in fact stimulate photosynthesis by zooxanthellae, but it is accompanied by a decrease rather than an increase in calcification [Langdon and Atkinson, 2005]. This may be due to competition between zooxanthellae and the host for the same internal pool of dissolved inorganic carbon [Marubini and Davies, 1996; Cruz-Pinon et al., 2003].
2.6.3. Seawater carbonate chemistry is the only variable that affects calcification
Although carbonate saturation state is a likely control on marine biocalcification, at least in some taxa, it is not the only variable that affects calcification rates in corals and other reef builders. In particular, two other variables that are known to affect calcification rates are temperature [Coles and Jokiel, 1978; Houck et al., 1987; Reynaud et al., 2003; Marshall and Clode, 2004], and light [Barnes, 1982; Chalker and Taylor, 1978; Marubini et al., 2001]. These variables help define the “optimum niche” for coral calcification, and their combined effects are addressed in Section 3.3.
Funny how all-or-nothing “misperceptions” can go both ways.
brandonrgates,
What’s your point?
Are you disagreeing with Jim Steele? If so, why not just say so, and provide some reasons for your thoughts.
Warmist tactics of deny, divert and confuse, don’t work as well as they used to. As you said, oops!
Cheers.
Read more, comment less. (H/T Steven Mosher)
Mike Flynn,
Start by comparing what Steele wrote about his citations vs. what they actually say.
I can’t do your thinking for you, Flynn.
MIke, I do not respond to Brandon Gates anymore and told him so. Last year at WUWT we engaged in a very civil discussion and I was very professional and provided him with all the evidence and links he requested. He then went over to Slandering Sou’s trying to smear me by portraying the interaction with nothing but ugly falsehoods. He is insincere not to be trusted.
Jim Steele,
Yes, you very civilly answered my questions and provided evidence up to the point that my line of enquiry began to expose the flaws in your argument. After which you professionally changed the subject. Here’s yet another example of you running a similar script:
There’s no need to trust my sincerity, Jim. How you have misrepresented the sources you cite in the OP is evident to any who take the time to make the comparison. Throwing rocks at the bearer of bad news does not diminish the bad news. Such behavior also doesn’t warrant my respect, politeness or civility.
You have only smeared yourself here, and there is no slander for calling lies out for what they are — wherever I choose to do it.
Good evening.
brandonrgates: There’s no need to trust my sincerity, Jim. How you have misrepresented the sources you cite in the OP is evident to any who take the time to make the comparison.
You presented very little. How about a few bullet points highlighting the specific points on which you think Jim Steel to have made misrepresentations? Or must we take it on faith that if we read all the references of Steele’s article we’ll assuredly agree with you? Claims like that have often been made here, with not a really good track record. This is a good place for point counterpoint counterpoint, etc.
FWIW, Steele’s presentation accords well with the other stuff that I have read in the peer-reviewed literature, granted not a great deal.
Start by comparing what Steele wrote about his citations vs. what they actually say.
“Likewise species inhabiting our warmest waters have undergone the fewest episodes of severe coral bleaching.”
The warmest spots have changed the least (your point apparently) – but they are still the warmest. His point was that it is variability not peak temperature that causes bleaching. Your quote did nothing to support a different conclusion.
BG 0, JS 1
“Studies of 7000-year-old fossil coral reefs in the South China Sea revealed high coral mortality every 50 years due to winter cooling events.”
Link supports his statement.
BG 0, JS 2
“Accordingly research has estimated that during the cold nadir of each ice age, coral reef extent was reduced by 80% and carbonate production was reduced by 73% relative to today.”
Can’t really award a point. Your quote and statement didn’t directly address his point. I’m sure you had a point – you simply didn’t express it or support it. The research says warmer is faster and the deepwater (cold) corals grow an order of magnitude slower. Research showing colder temperatures don’t inhabit calcification in tropical corals or better yet speed it up would have been relevant. You didn’t provide that..
PA,
It’s the *rate* of change in temperature, *among other things*, that coral experts are worried about.
No, not really: As their evolutionary history would predict, today’s greatest concentrations and greatest diversity of corals are found in the earth’s persistently warmer waters, like the Indo-Pacific Warm Pool. Likewise species inhabiting our warmest waters have undergone the fewest episodes of severe coral bleaching. Given their evolutionary history, coral’s greatest achievement has been enduring bouts of sustained climate cooling and rapid temperature swings.
His description is almost completely inconsistent with what the citation itself says. He’s also implying that higher absolute temperatures tend to be better for corals. Literature he cites suggests 16 to 18 °C *wintertime* temperatures are the lower bound. Further down he says this:
As the last ice age ended, coral expanded their range with warming temperatures. At the peak of the Holocene Optimum 10,000 years BP (Before Present), coral adapted to tropical ocean temperatures in the heart of the Coral Triangle were 2.1 °C warmer than today.
Only the first reference he gives leads to the full paper, and is dedicated to corals. The second one only goes to an abstract, which doesn’t mention corals at all. Maybe they’re mentioned in the body of the paper, who knows, he doesn’t provide a quote. Anyway, read the first paper; there’s a lot more to it than just absolute temperature of the water.
That fact does not rebut my argument: When most researchers start predicting an increased frequency of cold events, that will be especially salient information to consider.
That cold temperatures stress corals more than warm says zippo about whether corals can tolerate more warming at a unusually high rate in addition to falling ocean pH and any number of other human influences. Never mind the fact that nobody sane is worried about *falling* ocean temperatures. In sum, his argument here is a red herring. It matters not a whit that the citation is sound, how he’s applied it is the problem.
I quoted the title of the paper: Modeled estimates of global reef habitat and carbonate production since the last glacial maximum.
He only likes models when they support the point he’s trying to make. That’s my *opinion*.
Because that isn’t my argument. I’m happy to sum it up for you though: He’s over-simplified the argument down to cold = bad, hot = good, while largely ignoring several other influences contributing to thriving, healthy coral formations. He’s also peddling the notion that coral bleaching events are an adaptation mechanism whereas they’re more of a stress response which is neither infinitely sustainable, nor optimal to a reef’s health.
matthewrmarler,
I have this thing about not doing *all* of other’s homework for them.
Pretty much everything you need to make your own evaluation is right here on this webpage, otherwise Google is your friend. And no, I don’t expect you or most denizens here to assuredly agree with me. Ever. No matter how many bullet points I use.
Which is exactly what my first post in this thread did.
Not everything he writes in this article is out of accord with things I have read elsewhere in literature. It’s the conclusions he draws, or implies, which don’t follow from the citations — along with the few glaring discrepancies — I’m looking at.
brandonrgates | May 25, 2016 at 2:03 am |
His description is almost completely inconsistent with what the citation itself says. He’s also implying that higher absolute temperatures tend to be better for corals. Literature he cites suggests 16 to 18 °C *wintertime* temperatures are the lower bound.
Huh? The Solomon Islands has 537 and Raja Ampat has 534 species of coral. Implying that the coral are near a high temperature limit would supported by fewer species in the hottest spots not the planet’s personal best. Further reef corals grow around geothermal vents. Claims that high temperatures are an issue for reef corals should be viewed with disbelief.
You have done nothing to undermine his point that it is temperature variation (weather) and not long term temperature (climate) that is causing bleaching. Further the reloading with different zooxanthellae than were expelled makes it look like bleaching is a natural adaptation mechanism.
Finally, if variation is the issue in a period of warming temperatures there will be more warm bleaching events than cold bleaching events. When things start cooling it will go the other direction.
Mike Flynn,
I agree.
As an engineering major I was required to take a course in technical writing. In these classes they teach you to tell the reader what you are going to say, say what you want to say, and then conclude by telling the reader what you said.
All this is to be done in the plainest prose possible.
The climatariat doesn’t use this sort of language though. The language they use is what George Orwell in Politics and the English Language called “political language,” which “is designed to make lies sound truthful and murder respectable, and to give an appearance of solidity to pure wind.”
brandonrgates: It’s the *rate* of change in temperature, *among other things*, that coral experts are worried about.
That’s a “worry” — where is the evidence that Steele misrepresented any publication?
brandonrgates: I have this thing about not doing *all* of other’s homework for them.
I have this thing about discovering that people’s claims are not backed up by their links and citations — and then they either change their ground or insult me. You claimed that Steele wrote falsehoods and misrepresentations. Make the case so we can evaluate it. What you have written so far does not support your assertions.
I think you are either ignorant, lying, or delusional. If you write your points and link to or quote specific paragraphs, either you’ll show me wrong (it has happened), or help me evaluate which.
brandonrgates: It’s the conclusions he draws, or implies, which don’t follow from the citations — along with the few glaring discrepancies — I’m looking at.
That’s where your guidance on exactly where he went astray, and where his citations of evidence are accurate, and where you think his conclusions are unsupported would be most helpful.
I had an interchange some years ago at ClimateEtc in which a writer asserted that his case was made in a chapter of Pierrehumbert’s book “Principles of Planetary Climate”, which in fact I had read. I reread the chapter, and did not find his case supported, but inasmuch as he was unwilling to supply page and paragraphs, I could not tell for sure exactly what he had misunderstood, but he had clearly misunderstood it. We have been exposed to writers who have misunderstood the Feynman Lectures or the laws of thermodynamics. Some writers just make stuff up out of nowhere. So make your case and let us evaluate it, instead of saying that it’s all out there somewhere.
Some of the readers here, especially “lurkers” who never or seldom post, are sharp and open-minded. We hear from them now and again. Write for them.
PA,
Claims that some coral species thrive near geothermal vents means that thousands of species of coral living in typically cooler waters would do just fine next to a geothermal vent should be viewed as a basic failure to understand how species adapt to local conditions. Or maybe I should say, a basic failure to understand how species often do NOT adapt to local *changing* conditions. And as I have said several times, the *rate* of *change* in temperature isn’t the only factor.
Sweeping claims about thousands of species of corals living widely varying conditions being able to handle pretty much any change imaginable are the ones that should be viewed with skepticism bordering on disbelief.
MIke and PA, Thank you for pointing out Brandon’s dishonest and meaningless attacks. Sadly that is what he always does.
brandonrgates: Sweeping claims about thousands of species of corals living widely varying conditions being able to handle pretty much any change imaginable are the ones that should be viewed with skepticism bordering on disbelief.
On the other hand, the adaptability of large populations of small rapidly-reproducing creatures to changes in the chemical compositions of their environments has been widely demonstrated, in the form of acquired resistance of target organisms to pesticides, herbicides, fungicides, and antibiotics. All the evidence on adaptability supports the hypothesis that corals will adapt to changes in the ocean that are small relative to the interannual variability. If you have some evidence that corals are in fact not adaptable, evidence not “worry”, be sure to direct us to it.
Jim Steele said:
+1
matthewrmarler: “I think you are either ignorant, lying, or delusional.”
How about all three?
And disingenuous with it.
brgates, ” not sure what you mean with your first oops. The south china sea is in the middle of the indo pacific warm pool region but because of its currents and opening in the north east to deep water and colder more northerly waters, has a lot more temperature and nutrient variability than the “warm pool” proper, so the SCS has a stronger indication of the 50-60 year cycle that impacts most of the northern hemisphere.
Yes, I saw all his excerpts as supporting the idea that corals are complex and very adaptable and affected by much more than temperature. There’s no reason to believe the global warming will increase variability and increasingly harm corals. It doesn’t seem plausible that GHG concentrations and warming will adversely affect the ocean biosphere. Unless some wealthy evil genius finds and funds away to turn rock into CO2, I don’t think we can increase GHG concentrations very fast. In fact, I’m very concerned that we won’t be able to maintain adequate levels to keep all the areas that are become agriculturally productive going in the coming decades. The more the biosphere grows, the more it pulls CO2 out of the air, we aren’t likely to be able to extract fossils fuels fast enough to sustain increasing CO2 levels very much longer.
I think Brandon has missed the point. It’s not warm:good::cold:bad,, it’s that warm/=bad, it just is.
Even the desert ocean is full of life. The areas most likely to be affected acidify don’t seem to be much affected. The SH CO2 doesn’t increase as fast as the NH. There’s a lot to be inferred from that that blows up worries about CO2 increases and implications of GH effect.
Nice finds, thanks.
brandonrgates: Although carbonate saturation state is a likely control on marine biocalcification, at least in some taxa, it is not the only variable that affects calcification rates in corals and other reef builders. In particular, two other variables that are known to affect calcification rates are temperature [Coles and Jokiel, 1978; Houck et al., 1987; Reynaud et al., 2003; Marshall and Clode, 2004], and light [Barnes, 1982; Chalker and Taylor, 1978; Marubini et al., 2001]. These variables help define the “optimum niche” for coral calcification, and their combined effects are addressed in Section 3.3.
Of Steele’s points, which one does that contradict? That corals have survived great temperature swings? That corals can adapt to changing environments? That corals originated in warmer waters than what they now inhabit or will inhabit with a planet warming about 1C per century?
matthewrmarler,
Optimum niche and combined effects contradict the notion that because some corals do well in the warmest waters on the planet means that all corals would do well in a rapid warming regime. Or that some corals survived rapid temperature cycles in the past means that all present species would fare as well.
It’s the extrapolation from anecdote that is one of the main problems with this article.
Mathew, Thank you for revealing Brandon Gates irrelevant accusations. He is just one of Slandering Sou’s attack dogs trying to shoot the messenger and belittle anything that is does not promote climate change gloom and doom. Dont hold your breath waiting for an honest reply. He will obfuscate ad nauseum. He obviously lacks a scientific background because he attacks the essay simply for detailing the conditions within which coral evolved. But that is standard procedure for scientists. He even confuses peer reviewed literature suggesting it is anecdotal. But perhaps he does not believe in evolution.
He often creates false straw man arguments, which are the only ones he can cogently attack. For example no one has argued that just because these coral evolved in warm waters, or that they are now restricted to warm waters that it means they will survive an abrupt temperature change like an E. Nino. It simply means coral have survived warmer conditions than today and thus possess mechanisms that will allow them to adapt to any warming in the near future.
Often he does not understand what he is attacking.He try to suggest my link to the Kleypas paper stating that most coral in the warmest waters have not bleached. The paper mentions the ocean thermostat hypothesis. Because this essay was about adaptive bleaching I did not dwell on that aspect of the paper, but that paper rightfully suggest there is even less reason to worry about global warming because negative feedbacks prevent oceans from warming beyond a certain plateau.
The whole point of the essay was to detail emerging science revealing how coral can rapidly adapt to changing conditions, whether from warming or cooling, by shifting and shuffling their symbionts. The evidence for this is undeniable and the scientific literature abounds with evidence. It doesnt mean every colony or every reef will not endure mortalities. It simply means coral have evolved a marvelous system of adaptation and resilience to recover. Its perplexing that such good news does not incite awe and appreciation for the natural world. Instead Brandon and his ilk try to obscure these new scientific findings because they contradict his gloom and doom beliefs
brandonrgates: Optimum niche and combined effects contradict the notion that because some corals do well in the warmest waters on the planet means that all corals would do well in a rapid warming regime. Or that some corals survived rapid temperature cycles in the past means that all present species would fare as well.
If you have some countering evidence, do be sure to link to it.
Meanwhile, all the evidence presented supports the claim of adaptability.
Jim Steele,
No. I characterized your selective use of peer reviewed literature as an extrapolation from anecdote.
And perhaps you’re making things up. Any port in a storm, as they say.
It’s a basic tenet of biology that all species have adaptation mechanisms. What you leave out above is time — few species are capable of “instant” adaptation. Absolute temperature is not necessarily the deciding factor, but rate of temperature change. You also leave out that temperature isn’t the only environmental factor. Your conclusion that corals will be able adapt to any warming in the near future on the basis of such incomplete information and sloppy reasoning is risible.
matthewrmarler,
Not to “any warming in the near future” it doesn’t. Such a confident and unqualified statement warrants nothing less than instant dismissal.
matthewrmarler,
It’s really not my job to spoonfeed you that which you can Google for yourself. But what the heck, here’s a freebie:
Pandolfi et al (2003):
Degradation of coral reef ecosystems began centuries ago, but there is no global summary of the magnitude of change. We compiled records, extending back thousands of years, of the status and trends of seven major guilds of carnivores, herbivores, and architectural species from 14 regions. Large animals declined before small animals and architectural species, and Atlantic reefs declined before reefs in the Red Sea and Australia, but the trajectories of decline were markedly similar worldwide. All reefs were substantially degraded long before outbreaks of coral disease and bleaching. Regardless of these new threats, reefs will not survive without immediate protection from human exploitation over large spatial scales.
Coral reefs and associated tropical nearshore ecosystems have suffered massive, long-term decline in abundance, diversity, and habitat structure due to overfishing and pollution (1–7). These losses were more recently compounded by substantial mortality due to disease and coral bleaching (8–12). Although much longer records exist for some coral (13) and commercially important fisheries species (2,3), detailed ecological descriptions of reef ecosystems are less than 50 years old (14,15). The long-term historic sequence of ecosystem decline is unknown for any reef, thereby obscuring the potential linkage and interdependence of the different responsible factors that must be unraveled for successful restoration and management.
[…]
Historical trajectories of reef degradation provide a powerful tool to explain global patterns and causes of ecosystem collapse, as well as to predict future ecosystem states, allowing managers to anticipate ecosystem decline through an understanding of the sequence of species and habitat loss. Management options will vary among regions, but there must be a common goal of reversing common trajectories of degradation. The maintenance of the status quo within partially protected areas such as the Great Barrier Reef is at best a weak goal for management, which should strive instead for restoring the reefs that are clearly far from pristine. Regardless of the severity of increasing threats from pollution, disease, and coral bleaching, our results demonstrate that coral reef ecosystems will not survive for more than a few decades unless they are promptly and massively protected from human exploitation.
See again: it’s not all about absolute temperature.
matthewrmarler,
I’ll spot you one more. More recent, less alarming to my lay eyes, Descombes et al. (2015):
Abstract
Coral bleaching events threaten coral reef habitats globally and cause severe declines of local biodiversity and productivity. Related to high sea surface temperatures (SST), bleaching events are expected to increase as a consequence of future global warming. However, response to climate change is still uncertain as future low-latitude climatic conditions have no present-day analogue. Sea surface temperatures during the Eocene epoch were warmer than forecasted changes for the coming century, and distributions of corals during the Eocene may help to inform models forecasting the future of coral reefs. We coupled contemporary and Eocene coral occurrences with information on their respective climatic conditions to model the thermal niche of coral reefs and its potential response to projected climate change. We found that under the RCP8.5 climate change scenario, the global suitability for coral reefs may increase up to 16% by 2100, mostly due to improved suitability of higher latitudes. In contrast, in its current range, coral reef suitability may decrease up to 46% by 2100. Reduction in thermal suitability will be most severe in biodiversity hotspots, especially in the Indo-Australian Archipelago. Our results suggest that many contemporary hotspots for coral reefs, including those that have been refugia in the past, spatially mismatch with future suitable areas for coral reefs posing challenges to conservation actions under climate change.
brandonrgates,
With respect, the paper you liked to doesn’t appear to mention
temperature at all. Another Warmist red herring?
It’s a fairly awful paper, with the usual wild claims of coral reef ecosystems not surviving for more than a few decades without massive injection of Government funds (in practice). Pretty much like alarmist climate change predictions of imminent doom. An example of trend projection masquerading as science.
Species appear, species disappear. No more trilobites, stegosaurs or even woolly mammoths to be seen. So sad, too bad. Antarctica no longer maintains the rich flora and fauna it once did.
Corals? Seem to have adapted to most of the oceans, from hot to very cold. Without some reasonable anti-evolutionary theory, I assume that adaptation by evolution will continue, as it seems to have so far.
Increased temperatures in parts of the globe may have nothing to do with CO2, as there are a few other explanations which don’t require the greenhouse effect leap of faith. I prefer facts to faith, as far as science is concerned. I assume you do too, but you haven’t presented many so far.
Keep trying with the Warmist deny, divert and confuse. It might start working again.
Cheers.
Jim Steele said:
That’s because ‘Brandon and his ilk’ are not interested in the way the world is, they’re only interested in the way they believe the world ought to be.
That’s why what ‘Brandon and his ilk’ practice is not science, but religion. It may be secular stealth religion, but it is, nevertheless, religion.
The thing that causes ‘Brandon and his ilk’ the most gloom and doom is the fact that their gloom and doom predictions have failed to come to pass.
brandonrgates said:
http://www.faillol.com/wp-content/uploads/2009/11/jail-fail.jpg
One reason I refuse to respond directly to Brandon Gates is because the fails to read or fails to comprehend what was written.
For instance he makes the insane accusation that “What you leave out above is time — few species are capable of “instant” adaptation. ”
But the whole essay is about the speed with which corals adapt and the revelation the coral are one of species that are capable of rapid adaptation. Perhaps Brandon was upset and couldn’t get past the paragraphs that coral evolved and survived much warmer water.
Brandon also repeats the falsehood, “You also leave out that temperature isn’t the only environmental factor.”
FIrst it those attributing blame to global warming for bleaching that maintain that myopic view. In contrast I wrote,
“In contrast to researchers like Hoegh-Guldberg who emphasizes coral bleaching as a deadly product of global warming, bleaching is a visible stage in a complex set of acclimation mechanisms during which coral expel, shift and shuffle their symbionts, seeking the most beneficial partnership possible. Bleaching can be induced by stressful interactions between temperatures, disease, heavy rains, high irradiance from clear skies and competition with seaweeds. Indeed abrupt warm water events like El Nino have induced widespread bleaching and high mortality. But cold winters or La Nina induced upwelling of colder waters have also induced bleaching. ”
There was a whole subsection discussing other factors titled
2. Bleaching Causes the Least Mortality
Again it appears Brandon was so stressed by corals’ evolutionary history he never got past the first paragraphs. Or perhaps Brandon is blindly criticzing the wrong people. Whatever the case, his rants suggest a clear break with reality.
Jim Steele,
I don’t get into the nuts and bolts of many of these debates, such as this one, because I don’t know enough about the subject to take sides.
What I do look for, however, instead of content is the form the debate takes.
For instance, Brandon argues that, “It’s really not my job to spoonfeed you that which you can Google for yourself.”
That is an epic fail.
When someone makes a claim, it is incumbent upon them to marshall the evidence to back it up, not stomp their feet and say that, “If you’ll use the Google search engine you can find evidence to back up my claim.”
In short, what we get from Brandon is an orgy of rhetological fallacies. It’s all rhetoric and paralogisms, with a marked dearth of empirical evidence. There is no substance. It’s like boxing with a ghost. He’s like a strawman incarnate.
Well, the problem is the quoted studies are basically op ed pieces.
There have to be reefs in the Solomons or elsewhere in the hot zone where scientists could look at reefs that are just affected by temperature.
What do these reefs look like?
Perhaps some hard numbers?
The quotes from the studies are just hand wringing.. Man has been bad. He has overfished,, polluted, and diseased them (somehow). And now he is cimating them too!!!
Use of RCP8.5 for anything other than to wipe yourself is inexcusable. President Trump should have RCP8.5 removed from the scenarios because it is lacking in reality, and the projected CO2 levels are diverging so rapidly from reality so fast that it will soon be even more irrelevant than it is now (Irrelevanter?)..
And then as a parting shot the quotes mention that the range of corals is expanding northward.
It sort of is what it is. Are corals going to die out? No. Are some of them going to have to move? Yeah, probably.. Corals do best in nutrient deficient water that doesn’t support starfish. Unless you introduce a virus ( or some other solution) and kill off the starfish that isn’t going to change.
brandonrgates: Not to “any warming in the near future” it doesn’t. Such a confident and unqualified statement warrants nothing less than instant dismissal.
Who says its confident and unqualified? It’s supported by the evidence.
How much of random variation and natural selection are you disputing here? There is no such thing as an “optimum” environment: within any environment, the progeny in each generation show much variation, and within each species except a few large species like humans nearly all of the progeny die before they reproduce. Among each species of coral in each environment, some progeny will survive better at a higher temp, some better at a lower temp.
Coral reefs and associated tropical nearshore ecosystems have suffered massive, long-term decline in abundance, diversity, and habitat structure due to overfishing and pollution (1–7).
but that says nothing about future warming. Nobody has written that “it is all about temperature”, but “global warming” is about temperature, and that reference (of which I took only a short quote) was incidental to warming.
Here is another: Related to high sea surface temperatures (SST), bleaching events are expected to increase as a consequence of future global warming. However, response to climate change is still uncertain as future low-latitude climatic conditions have no present-day analogue
That expectation is at least is testable, but takes no account of random variation and natural selection (or of recorded change), which I repeat are among the most reliably demonstrated phenomena in nature. It has been well demonstrated that expert opinions in the AGW debate sometimes go beyond what can be supported by evidence (there was even a paper published calling it a new paragidm), and are sometimes (as here) in defiance of evidence. Granted that the future is not known, we should at least give more weight to evidence than to unsupported expectation.
It isn’t your “joob” to show that your critique of Steele’s essay has merit, but this is an opportunity to support your critique, and so far you are blowing it. You write about other problems than warming, when warming is the problem asserted by CO2-AGW theory, and you cite expectations that are discordant with published evidence. There are people at ClimateEtc who regularly write in support of the CO2-AGW consensus, so you need not fear an “echo chamber” of “deniers”. Everyone who has evidence in support of your critique will bring it in.
matthewrmarler,
I do. I should have written overconfident and unqualified.
What’s supported by evidence is that *some* corals have been able to survive rapid temperature changes, and that corals generally prefer warmer water to cooler. And every coral expert in the world will tell you that bleaching is an adaptation that increases survivability when corals are stressed for any number of different reasons.
What is NOT supported by evidence is Steele’s conclusion: It simply means coral have survived warmer conditions than today and thus possess mechanisms that will allow them to adapt to any warming in the near future.
That’s a ridiculous prediction on the face of it.
None. I’m disputing the prediction that *any* amount of warming in the near future is something that corals will survive because … adaptation.
Let’s rewind to that citation: These variables help define the “optimum niche” for coral calcification, and their combined effects are addressed in Section 3.3.
“Optimum niche” is in “scare quotes” which means in this case that it’s being used as a “figure of speech”. The authors are prevailing on the assumption that anyone reading that particular text should have by then obtained a basic knowledge of adaptation and/or speciation such that they recognize evolutionary processes don’t result in optimal adaptations, only adaptations which favor fitness of certain individuals to a particular ecological niche over other individuals.
Think of it this way. In a group of humans running from a bear, nobody can run faster than the bear. Everyone only need be able to run faster than the slowest individual in the group. The optimal adaptation is that every individual can run faster than the bear.
Sure. In many aquatic species, especially those who don’t care for their young, the reproductive strategy is spamming the water with gametes. Which is what corals do, as well as being capable of asexual reproduction. That still does not ensure that corals will be able to handle any amount of near term warming.
Among each species of coral in each environment, some progeny will survive better at a higher temp, some better at a lower temp.
Plausibly true. I’m not a coral expert. Not even close. If you were a coral expert, I’d be asking you what the distribution and range of temperatures were, how that varies among the thousands of species of corals, and what factors *other* than temperature might limit their adaptability to *any* near term warming.
I keep saying this isn’t all about temperature.
Quoting Steele again: It simply means coral have survived warmer conditions than today and thus possess mechanisms that will allow them to adapt to any warming in the near future.
*Any* warming in the near future.
And yet Steele is (over)confident that corals will be able to adapt to any near future warming despite expressed uncertainty in literature, even the stuff he’s cited.
One of the most uncontroversial conclusions of biology is that mass extinctions happen when there are rapid environmental/ecological changes.
It has been well demonstrated that inexpert opinions in the AGW debate sometimes go beyond what can be supported by reason.
Yeah, that’s about as good a summary of the sum total of my argument against Steele’s essay.
One way to reduce uncertainty in future expectations is to change as few variables in the system as possible. Another way to reduce uncertainty in a hurry is to say hell with it, damn the torpedoes, full speed ahead.
Jim Steele,
It simply means coral have survived warmer conditions than today and thus possess mechanisms that will allow them to adapt to any warming in the near future.
I think it’s fair to suppose one reason you “refuse to respond directly” to me is that I have this nasty habit of spotting your self-contradictions. No particular scientific expertise required to do that, I might add.
Glenn Stehle,
lol. Ok, sure. It’s my job to teach this entire forum about corals. One thing I do know for sure, y’all aren’t paying me enough to do your homework for you.
it is fortunate that BG is not a medical examiner.
He would do an autopsy on someone who was poisoned, choked, had the plague, and was half eaten, then conclude they died from hyperthermia.
PA says “it is fortunate that BG is not a medical examiner.
He would do an autopsy on someone who was poisoned, choked, had the plague, and was half eaten, then conclude they died from hyperthermia.”
ROTFLMAO That is exactly Brandon Gates. He is a Slandering Sou acolyte. Their trademark is to ignore all the evidence, find two sentences that they can dishonestly juxtapose, and harp on it in order to hijack a thread. They never engage in sincere and honest discussion.
My essay has two sections talking about factors other than temperature. But Brandon Gates ignores all that and dishonestly uses a quote out of text to portray himself as the only one looking at all the factors. He oozes dishonesty.
The quote, “It simply means coral have survived warmer conditions than today and thus possess mechanisms that will allow them to adapt to any warming in the near future” is only one of many points. And indeed if he understood evolution, and if he understood there is a pool of “pre-adapted” symbionts that can be shuffled and shifted, their adaptive mechanisms virtually guarantee that coral will be able to adapt to the same higher temperatures they survived during the Holocene Optimum. There is absolutely no scientific reason to argue they won’t. And Brandon that doesnt mean I think a coral will survive dynamite fishing. LOL
Brando Gates is such a legend in is own mind, he believes he sees contradictions that do not exist. He has absolutely no understanding about coral biology and adaptive mechanisms. His only expertise is word-smithing BS.
PA,
What about the studies Steele cited?
That’s what controlled laboratory conditions are for.
Yeah, that would be nice wouldn’t it. Let’s not forget the time component.
I see, so it’s absolutely impossible that pollution, overfishing and disease could possibly ever effect corals. Good to know, I’m so relieved.
By the way, I have several metric tonnes of toxic waste I’ve been meaning to get rid of, may I use your front yard? It shouldn’t negatively affect any humans that might happen by, we’re a resilient species known to have survived many prior instances of adverse environmental conditions.
It’s a moot point, really. According to Steele, corals will be able to handle any near term temperature change nature can throw at them.
PA,
True. First decomp case and I’d be out. Hospital morgues are routinely less smelly.
What this has to do with corals, I’ve no idea, but do please carry on.
Jim Steele,
And your comments contain one sentence which makes it all for nought: It simply means coral have survived warmer conditions than today and thus possess mechanisms that will allow them to adapt to any warming in the near future.
*Any* warming, Steele. Nothing else matters. Not my fault that you’re so sloppy.
Pre-adaptation!!! Ok ok, peppered moth, you’re not talking total nonsense here. And let it not be said I eschew punctuated equilibrium for strict gradualism or vice versa. But virtual guarantees because … pre-adaptation? You’re killing me here.
How long does it take these pre-adapted symbionts to be shuffled and shifted for a given set of environmental changes? Obviously not immediate, otherwise there wouldn’t be sustained bleaching episodes.
Kim Cobb of Georgia Tech:
http://www.climatecentral.org/news/dead-coral-reefs-climate-adaptation-20241
Unprecedented temperatures yet they are confident of recovery it seems to me.
“We’re really interested in understanding who is coming back and how well are they able to thrive and and what the role of these resilient little miracle corals is. This is basically adaptation on speed.”
Agile coral. Those crazy crazy coral. Thrive? Things will be Okay.
PA writes Well, the problem is the quoted studies are basically op ed pieces.
Brandon Gates avoids the issue with another dishonest distraction “What about the studies Steele cited?”
Out of about 50 links about 46 were to peer reviewed literature. I linked to my essay on the Antarctic Refrigeration effect that linked to 20 peer reviewed studies regards the Cenozoic cooling trend. Another to a NOAA report, and 2 to wikipedia for islnd descriptions. Every other link should have been to peer reviewed science publications.
So is Brandon engaging in his standard dishonest distractions? Perhaps he can point out all my “op eds”
Jim Steele,
And I linked to two peer-reviewed publications and one galley proof of a textbook. It’s not your references that are the problem, it’s your home-grown prediction that corals will be able to handle any amount of warming in the near future that’s at issue.
Do please continue tap dancing around the problem with your main thesis while pretending I’m the one who is distracted.
Ragnaar,
Yes, that’s what Steele is selling: any amount of warming in the near future will be a-ok. Let’s read what the article actually says:
There’s a Hopeful Message Hidden in These Dead Reefs
Published: April 13th, 2016
By Brian Kahn
“It was a gut punch.”
That’s how Kim Cobb, a scientist at Georgia Tech, described what she saw on a recent dive on the reefs surrounding Christmas Island. She’s studied them for the better part of two decades and what she saw this April was something she had never seen before.
Once vibrant reefs, some of the most pristine on the planet, were almost completely dead. Brownish red algae was draped across them like a funeral shroud. Hot water brought on by one of the strongest El Niños on record — layered on top of climate change-driven warming — was simply too much for the reefs to take.
There were exceptions, though — tiny pieces of baby coral that are beating the odds. And those exceptions could give scientists insight into how to make sure climate change doesn’t completely destroy reefs around the world.
“This is a race against time for coral reef biologists around the world to understand what makes certain corals able to survive,” Julia Baum, a coral researcher at the University of Victoria, said. “If we can understand that, then we can start to develop targeted solutions to preserve corals throughout the next century of climate change.”
Right. So this reef all but dead since the 1998 El Nino, and it’s now starting to show some signs of recovery. Other coral scientists are interested because that could be key to understanding something which presently isn’t: what are the factors which maximize survivability and how might we enhance their native ability to do so. Notice all the qualifiers and conditionals. Real scientists are cautious about making too strong of conclusions on the basis of limited evidence … which this is: it’s one reef. It’s hopeful, and potentially good news, but not a lock, and looks to still require our intervention IF reef recoveries/preservation in other areas are to be successful.
These are very different statements from Steele’s virtual guarantee: pre-adapted symbionts will allow corals to handle any amount of near term future warming.
brandonrgates: It has been well demonstrated that inexpert opinions in the AGW debate sometimes go beyond what can be supported by reason.
True, but you are making that claim about Steele’s essay in particular, without basis.
And you don’t know that the progeny of corals show great variation, and the history of chemical alterations of environments of insects and bacteria is irrelevant, but you think that running from bears is relevant?
brandonrgates:
All but dead since the recent El Nino. The writer paints a dreary picture. That is contrasted with the Cobb quote. It works well I think.
matthewrmarler,
Rubbish. I have gone to some detail explaining how and why I think Steele’s central thesis fails. Since his argument has evolved from first posting, let’s review the progression:
Other coral experts have noted that coral reefs are already in decline, and have been for several centuries. They point to human overfishing, falling pH due to increased concentrations of atmospheric CO2 in addition to rising ocean temperatures as *combined* factors.
None of those other coral expert opinions necessarily falsify the adaptive Bleaching Hypothesis. It does however tend to challenge your unqualified and over-confident prediction that corals will be able to adapt to *any* near term temperature rise. Let’s look at the evolution and adaptability of your own narrative:
Notice that:
1) “hypothesis” has morphed into “virtual guarantee”
2) what causes most coral mortality is only alluded to in the OP, but Steele’s prediction is based only on one factor (temperature) and all but exclusively emphasizes recovery from bleaching events due to abrupt but not sustained temperature increases
3) he moves the goalposts from “will allow them to adapt to any warming in the near future” to “will be able to adapt to the same higher temperatures they survived during the Holocene Optimum”
So, overconfident and silently-changed predictions from a very limited set of factors known to literature. I’ve also previously documented how he misrepresents the literature he has cited.
That you dispute my conclusion that Steele’s essay is misleading does not mean that I have not provided any basis for making that claim.
I know that there are some 2,500 different species of coral, about a thousand of which are reef-builders. I also know that the geologic record clearly indicates that the chemical composition and other physical parameters of the biosphere (like temperature) have been anything but static. Since the planet is not presently sterile, that obviously implies that *all* life is resilient and adaptable to changing conditions.
None of that diminishes from what the geological record also shows that mass extinctions happen when there are *relatively* rapid changes to local environments. The species which do survive are the ones either best adapted to their local niche, are mobile enough to migrate to a more suitable environment, and/or those which a few fortuitously hardy individuals survive to repopulate their numbers. That’s not an exhaustive list; however, it does speak to the relevance of the Running from Bears metaphor.
In sum, adaptability and survivability as demonstrated in the past does not even virtually guarantee future survivability of any species, come what may. Far from it.
Continuing to strawman me is probably not your best tactic here. As such, do please continue doing it.
Ragnaar,
Cobb’s quote works well to say, “It’s not as bad as I thought.” Or, “There’s some hope we can do something to remedy what’s already begun.” Her quote doesn’t work well to suggest that everything will be a-ok come what may with little or no intervention on our part.
Basically, you may choose to ignore the already realized bad news in light of the *potential* good news, but that doesn’t make the bad news go away. To put it another way, there’s a difference between wishful thinking and rational hope, especially when the rational hope looks to be *contingent* on taking some set of actions … not all of which are presently well-understood.
Brandon Gates continues to his dishonest misrepresentations of the essay saying:
Gates suggest 1) “hypothesis” has morphed into “virtual guarantee”
Hypothesis remains as it always has and as with every hypothesis, its validity and predictions will be tested by future observations. That my comments suggest that the overwhelming evidence that has emerged virtually guarantees that past resilience and symbiont shuffling argues for success is no different than an IPCC “highly likely” argument. Gates is just trying to create a context to within which to insert his treademark enigrating remarks.
Gates outright lies again suggesting 2) what causes most coral mortality is only alluded to in the OP, but Steele’s prediction is based only on one factor (temperature) and all but exclusively emphasizes recovery from bleaching events due to abrupt but not sustained temperature increases
Gates continues to present this falsehood despite he was exposed for this falsehood earlier.
There is a subsection Multiple Causes of Bleaching
Gates further distorts “3) he moves the goalposts from “will allow them to adapt to any warming in the near future” to “will be able to adapt to the same higher temperatures they survived during the Holocene Optimum”
Another typical Gates distortions. Those quotes are simply one and the same argument. Gates trademark is to juxtapose sentences he doesnt understand and suggest they are contradictions or moving goal posts.
Does Gates have any integrity??
brandonrgates:
I may be more optimistic than you and quoted Cobb as I thought what she said supported optimism. I am more of a thrive than a mitigation person. While the El Nino event was tragic for some coral, other coral apparently will thrive and adapt. Oh and I see she worked ‘resilient’ into what she said too. Let’s assume that the normal state there is cool to average with the periodic El Nino ruining some corals week. Some of them seem to be ready for the non-regular El Nino warmth blast. Just a random thought, local sea levels tip them off as to when the next El Nino will happen. They are anti-fragile. I suppose I see what may not be there. The good news is life is resilient. I would suggest that’s true, because here we are in the Garden of Eden, some sins allowed.
Just some musings. Some coral have a stupid gene pool. It can’t adapt to change. The occasional El Nino improves the gene pools by taking the stupid coral out, leaving behind the a stronger gene pool of coral with smart gene pools. The recent El Nino lead the coral. It told the coral, it’s going to get warmer. Now adapt to it. Variation itself, leads the coral to where it needs to be to survive.
Jim Steele,
Hence it would be folly to make a “virtual guarantee” on the basis of a hypothesis.
To be consistent you must therefore accept all projections and predictions the IPCC considers “highly likely”. Somehow I doubt you do.
Let’s look again at what you have written:
In comments, you conclusions are that corals will either:
1) be able to adapt to *any* near term warming or
2) be able to adapt to warming up to at least those seen at the Holocene Optimum
You say in the case of (2) that this is a “virtual guarantee” based on the Adaptive Bleaching Hypothesis.
Yes I read that. I also read this statement in the OP: * Bleaching is not responsible for most coral mortality.
Your “virtual guarantee” of coral survivability ignores other causes which you yourself say are responsible for *most* coral mortality. Your temperature-centric guarantee doesn’t follow from its own premises, Jim. That’s not my issue, it’s yours.
Ragnaar,
I’m reading everything she says. It’s a *qualified* optimism, one which she says is *contingent* on:
1) Futher understanding what contributes to coral survivability and recovery
2) Developing targetted solutions to preserve corals over the next century of climate change
That’s already at odds with Steele’s conclusion that corals are virtually guaranteed to survive in temperatures up to the Holocene Optimum. It’s even further at odds with what her colleague Julia Baum said in the concluding paragraphs of the article:
The findings could help scientists create super corals that are adapted to future warmer waters. It’s a process called assisted evolution, and it’s highly controversial in the marine conservation field where protected areas are viewed as the gold standard for keeping ocean species safe from the ravages of climate change, fishing and other human actions. But after witnessing what happened at Christmas Island, Baum thinks it could be essential if we want coral in our future.
“Marine protected areas are not immune to climate change. Blind faith that they will protect coral from the types of hot water events we just saw is ludicrous,” she said.
The final sentence is particularly at odds with Steele’s virtual guarantees.
Let’s get something clear here: the biosphere doesn’t care about your personal philosophy or optimism any more than it cares about my implied pessimism. It only responds to the sum of its inputs, outputs and internal interactions, of which we are a part.
Not will, but may or might adapt. According to the article you cited, possibly not without active intervention on our part.
Some, not all. The question is how much is some. Let’s not assume we know that with any degree of certainty.
I don’t see how.
Just a WAG: I give them better odds than the megafauna species which perished around the time of the Holocene Optimum. Corals have made it through the Big Five major extinction events. It’s worth noting that they often didn’t reappear in the fossil record for hundreds of thousands to millions of years after having disappeared, in line with the severity of the events leading to global die-offs of many (most, in some cases) known aquatic and terrestrial species.
All sins have consequences. Major sins have major consequences. We don’t need every coral species to go extinct for it to be a Bad Thing for a lot of people.
I can’t force you to see what I see. All I can do is tell you what I do see. On corals what I see is that it looks bad, but there’s hope we can avoid the worst IF we take active measures to do something about it. I call that pragmatism, YMMV.
“It’s a *qualified* optimism, one which she says is *contingent* on…”
Yes it seems possible that Cobb is a doctor and needs to heal the sick patient. What I copied of what Cobb said was what I thought was glowing endorsement of what Steele is trying to write.
https://curryja.files.wordpress.com/2013/08/thrivability.gif
But no it’s not the whole story.
I agree with Steele that “…corals are virtually guaranteed to survive in temperatures up to the Holocene Optimum.”
Corals first formed about 240 million years ago. Call it a successful thing.
Baum is more left on this yes. Engineering corals or whatever the idea is like helping turtles cross the street. I’ve done that. Big 10 pound snapper.
Here’s what I linked to in summary:
1) Writer – It’s bad
2) First scientist – Amazing adaptation
3) Second scientist – We may have to operate on the patient
“Let’s get something clear here: the biosphere doesn’t care about your personal philosophy or optimism any more than it cares about my implied pessimism. It only responds to the sum of its inputs, outputs and internal interactions, of which we are a part.”
The biosphere cares little for what I do. I am still trying kill some carp to improve a lake, but I never seem to make progress with that. It’s been my observation or guess that life, in this case corals are less threatened by changing climate conditions than we give them credit for. They are complex and it may not reduce to Temperature times X equals the number of coral. But if it does we are back to sensitivity. What is X? And what temperature is ideal for its evolutionary success? Seem that would be a variable temperature. Perhaps we should be learning about ourselves from the coral and not trying to change them as Baum hints at. Insert Gaia argument here. We could know little of the outside world for decades lacking our smartphones and access to the internet on a Pacific paradise. We could still know an El Nino had happened. We would just ask the coral.
brandonrgates: In sum, adaptability and survivability as demonstrated in the past does not even virtually guarantee future survivability of any species, come what may. Far from it.
No one has written of “guarantees”: some experts fear the future (you have quoted some), despite loads of evidence of past survival of corals in situ, loads of evidence for random variation and natural selection generally, and loads of experience of humans trying unsuccessfully to eradicate large populations of small organisms by killing them with chemicals. If there is any reasonable expectation based on records and experience and well-tested theory (evolution by random variation and natural selection), aka an “induction” or “abduction” (so called by C. S. Peirce), it is that the corals will survive this temperature rise, which is small in comparison to past changes and interannual and regional variability.
I have read your interchanges with Steele, and I have to say that the main points of his essay have withstood your attempts at critique. I can’t see where anything that you wrote or quoted undermines what he wrote. You go off on tangents, invent exaggerations, ignore masses of evidence, and avoid answering direct questions. Where is the evidence that warming at current rates will harm the ocean’s corals?
brandonrgates: Pre-adaptation!!! Ok ok, peppered moth, you’re not talking total nonsense here. And let it not be said I eschew punctuated equilibrium for strict gradualism or vice versa. But virtual guarantees because … pre-adaptation? You’re killing me here.
How long does it take these pre-adapted symbionts to be shuffled and shifted for a given set of environmental changes? Obviously not immediate, otherwise there wouldn’t be sustained bleaching episodes.
Second point first, of course it is not “immediate”, but requires multiple generations.
“Pre-adaptation” merely means that among the progeny there are, by random variation, some who will reproduce better in the changed environment. In this case, it merely restates the case that some corals will reproduce faster in a warmer environment than most of their progenitors. As you noted below, some of the mechanisms are already known for corals.
Not necessarily. At least, not necessarily “multiple coral generations.” It is, in principle, possible for an individual coral to expel its symbiont load and rebuild it from surrounding samples, just as it’s possible for humans to expel their symbiont load (massive vomiting and diarrhea) and rebuild from surrounding samples. (In fact, bleaching in coral makes a fairly good analogy with such responses in humans.)
There are several other mechanisms that haven’t (AFAIK) seen much attention here.
For instance, symbiotic relationship behavior, in addition to being potentially genetic, could also potentially be epigenetic. My studies on that subject are most of a decade out of date, and focused primarily on DNA methylation, but even that has enormous promise for supporting short-term adaptation.
I see little sign of focused studies on this issue WRT coral bleaching recently (your mileage may vary). But even my decade-old understanding of DNA methylation suggests that evolved-in adaptive mechanisms could be present.
Normally, methylation patterns are heritable, although many gametes have specific mechanisms to reverse much of the parent-supplied load. But specific switches, set and/or reset by environmental factors, can have their settings inherited until/unless new environmental factors intervene, often many generations downstream.
Just remember that such epigenetic mechanisms have to have arisen by normal, Darwinian genetic evolution, and be supported by ongoing selection. Such mechanisms are complex, and will probably be lost if they don’t stay adaptive.
Jim –
You write…
And then you write:
But there is no reference for those “other researchers” and what “they argue,” Searching for the string in Google for what “they argue” that you quoted only returns a hit to your same article written here but posted elsewhere. Further, the hypertext link further up in the paragraph under “others have accused him” of persistently….” returns a hit to an abstract that doesn’t contain the quote, and the previous hyperlink connected to “popularizing worst case scenarios” is broken..
Could you provide better citations for what “they argue?”
I apologize for any trouble with my links. Most of the papers I download are via my university privileges so I avoid paywalls. But those links do not work for others. So I google the title, and use the first link provided. Sometimes it only provides the abstract but the full text is paywalled. So you will need to search for the title and find a free version or pay. I try to link to the papers from which I quote but sometimes I just miss one or lose while editing. Sorry for any inconvenience
1. The link to “popularizing worse case scenarios” should be
http://link.springer.com/article/10.1007%2Fs00338-008-0432-1
2. The link to …..They argue, “A view of… should be
http://gefcoral.org/Portals/25/BWG/EcologicalMonograph2007.pdf
There was a problem with how my formatting translatied to Dr. Curry’s site so I am not sure what other problems may have arisen. The article is also posted on my website http://landscapesandcycles.net/coral-bleaching-debate.html if that helps
Jim –
Can you reconcile the difference between this:
And this?:
1. It wasn’t a clear reference to Hoegh-Guldberg.
2. It wasn’t an “accusation,” in fact, it was a statement about the appropriateness of popularizing worst case scenarios.
And how about that “others have accused him of misunderstanding and misrepresenting the adaptive bleaching hypothesis.” ???
Can you give a specific reference? Who are the “others” that you’re referencing? It seems I don’t have a search feature available for PDF’s.. so page and paragraph/line numbers would be greatly appreciated.
Oh, and Jim.
No need to apologize about the links, it happens.
Jim –
Please note, from one of the links you provided (an article I found very interesting article, btw).
Do you disagree with any of the parts that I bolded? Is the underlying thesis that you argue in your essay is consistent with the quote I provided, in particular the parts I bolded?
No doubt, the authors of that article argue a thesis that is consistent with this characterization of yours:
But as with the excerpt I gave above, I wonder whether you think that this:
(particularly the part I bolded) is consistent with the thesis you present in this blog post.
Joshua ask “Can you reconcile the difference between….
1. It wasn’t a clear reference to Hoegh-Guldberg.
2. It wasn’t an “accusation,” in fact, it was a statement about the appropriateness of popularizing worst case scenarios.”
Joshua your quibbling is a bit suspect. The authors of the paper are clearly concerned with coral mortality and as their final paragraph explains they are concerned that when worst case climate scenarios get hyped in the media they can promote a sense of hopelessness in the public instead of encourage action that could promote resilience. They also rightfully argue if worst case scenarios are to be presented, then they those scenarios should not be based on untested assumptions. You can quibble with the word accuse, but when they identify who is disseminating problematic untested assumptions, they most often point to HoeghGuldberg papers.
For example, “Predictions that reefs will disappear as a result of global
warming are based, at least in part, on the assumption that corals are living close (within 1–2°C) to their maximum thermal limits (Hoegh-Guldberg 1999).”
They go on to discuss the wide range of temperature tolerances.
Regards the assumption coral can not adapt rapidly they wrote,
“That corals lack the capacity to adapt to projected rates of environmental change is a hypothesis based on three key assumptions: (1) that generation times are too long to allow for adaptation over the required timeframes (Hoegh-Guldberg 1999); (2) that the scale of dispersal is too large to allow for adaptation to local conditions (Potts 1984); and, (3) that there is insuYcient genetic diversity in existing symbionts and corals (Hoegh-Guldberg et al. 2007).”
This essay is about the growing support for the adaptive bleaching hypothesis and rapid adaptation due to symbiont shuffling and shifting. The evidence I presented here is why those authors challenge Hoegh-Guldberg’s assumption that coral can only adapt over long evolutionary time frames.
As I wrote bleaching is the least of coral reefs’ problems and only a minor cause of mortaity from which they typically recover from in less than a decade.There is a thin line between “promoting worst case scenarios” and fear mongering. When those scenarios are based on bad assumptions I suggest it fear mongering not good science. But Hoegh-Guldberg has hitched his fame and fund raising fortune to such scenarios, despite the emerging evidence of coral’s ability for rapid adaptation, he has dismissed the works of many experts that expose the errors of his assumptions.
Joshua ask “Can you reconcile the difference between….
..
1. It wasn’t a clear reference to Hoegh-Guldberg.
2. It wasn’t an “accusation,” in fact, it was a statement about the appropriateness of popularizing worst case scenarios.”
Joshua your quibbling is a bit suspect. The authors of the paper are clearly concerned with coral mortality and as their final paragraph explains they are concerned that when worst case climate scenarios get hyped in the media they can promote a sense of hopelessness in the public instead of encourage action that could promote resilience. They also rightfully argue if worst case scenarios are to be presented, then they those scenarios should not be based on untested assumptions. You can quibble with the word accuse, but when they identify who is disseminating problematic untested assumptions, they most often point to HoeghGuldberg papers.
For example, “Predictions that reefs will disappear as a result of global
warming are based, at least in part, on the assumption that corals are living close (within 1–2°C) to their maximum thermal limits (Hoegh-Guldberg 1999).”
They go on to discuss the wide range of temperature tolerances.
Regards the assumption coral can not adapt rapidly they wrote,
“That corals lack the capacity to adapt to projected rates of environmental change is a hypothesis based on three key assumptions: (1) that generation times are too long to allow for adaptation over the required timeframes (Hoegh-Guldberg 1999); (2) that the scale of dispersal is too large to allow for adaptation to local conditions (Potts 1984); and, (3) that there is insuYcient genetic diversity in existing symbionts and corals (Hoegh-Guldberg et al. 2007).”
This essay is about the growing support for the adaptive bleaching hypothesis and rapid adaptation due to symbiont shuffling and shifting. The evidence I presented here is why those authors challenge Hoegh-Guldberg’s assumption that coral can only adapt over long evolutionary time frames.
As I wrote bleaching is the least of coral reefs’ problems and only a minor cause of mortaity from which they typically recover from in less than a decade.There is a thin line between “promoting worst case scenarios” and fear mongering. When those scenarios are based on bad assumptions I suggest it fear mongering not good science. But Hoegh-Guldberg has hitched his fame and fund raising fortune to such scenarios, despite the emerging evidence of coral’s ability for rapid adaptation, he has dismissed the works of many experts that expose the errors of his assumptions.
Joshua, Clearly, by your selective bolding, you want to emphasize the point the warmer temperatures can cause bleaching. That is odd because that point has never been questioned and it is an issue I discussed. Coral’s evolutionary history and current geographic locations simply suggest coral are better adapted to warmer conditions, but they to have survived they have had to adapt to cooler conditions as well.
To repeat, the recent global bleaching events are typically the result of warm El Nino events, where warmer waters intrude into regions that are normally cooler. This essay is about how bleaching is part of coral adaptation mechanism, whether or not it caused by warming or cooling. Because coral attempt to maximize photosynthesis by acquiring the symbiont best adapted to their microclimate, coral engage in symbiont shuffling and shifting. Warm El Nino events are the primary cause of abrupt warming beyond normal limits and do result in bleaching. But bleaching occurred in the 1800s and early 1900s due to warming events relative cooler ocean temperatures. Bleaching happens no matter what the background ocean temperatures are. It is the rapid departure of temperatures over a period weeks and months that provoke bleaching, whether or not there has been natural or anthropogenic climate change, or no change at all. I have highlighted the fact the coral are very susceptible to cooling, because coral do not care what the models suggest, they must adapt to abrupt warming or cooling, because as we have observed cold winters and cool La NInas are also stressful.
In response to an El Nino driven event there is acquisition or survival of symbionts that are best adapted to the relatively warmer conditions. But post El Nino cooling often results in shifting to cooler adapted symbionts. Coral have no way of determining if a warm or cool event is a short term weather event like El Nino or long term change such as warming after the LIttle Ice Age. So bleaching and symbiont shuffling must respond to extremes above or below the average. In the warmest waters like the Western Warm Pool where El Ninos have less of an effect, there is less bleaching because there is less variability. The point of the essay is coral bleaching is a marvelous adaptation mechanism to deal with climate variability and has enabled coral to survive millions of years of dramatic change.
Jim –
==> “Joshua your quibbling is a bit suspect.”
Again, compare and contrast:
and
On the matter of “quibbling.”
I think that your characterization of what they said is quite different from what they’ve said. This is inherently a matter of interpretation. People, IMO, should be open to questions about situations where there is inherently some ambiguity.
It seems to me that the issue of “hyping” is a valid question, but that it’s extremely important to be careful when discussing what is and isn’t “hype,” and when discussing what scientists say. I don’t consider that to be a matter of “quibbling.”
There are two basic ways to investigate these issues. One is to conduct the science, the research itself.
Another is to characterize what the research shows. IMO, when characterizing what the research shows,it is important to be extremely careful to control for biases, in fact to bend over backwards to be sure to portray the full range of viewpoints – otherwise you run the risk of not accurately portraying what the science says and in the end, contributing to, and in some cases perpetuating, the gap between what scientists say and how the discussion plays out in among non-scientists.
Jim –
==> “Joshua, Clearly, by your selective bolding, you want to emphasize the point the warmer temperatures can cause bleaching.”
Not exactly. Let me repost what I bolded:
and
and
Now I asked you some direct questions about what I bolded. I think those questions make it quite clear what I was emphasizing by having bolded certain points. I notice that in responding you did not answer those questions.
Obviously, I can’t make you answer those questions, but if you are interested in addressing what I’m emphasizing, perhaps answering those questions would be the first place to start? If what I’m emphasizing wasn’t clear, I could attempt to elaborate, but I don’t know that there would be any point in doing so if, instead of even attempting to address my questions, you wrongly form impressions that you are “clear” about, and state them as fact.
Josh, You appear to be obsessing over a single sentence that a case can be made for worst case scenarios. Again we all agree that there is a place, but only if those scenarios are based on valid assumptions. But you are attempting to side step the meat of their paper in which they regards “predictions that functioning reef ecosystems will vanish within decades” by Hoegh Guldberg and his ilk
“The question is are they, like Cassandra, correct, or are these the false prophets that Grigg (1992) warned us of 15 years ago? Only time will tell, yet the premises behind some of the predictions can be examined,
and we argue that many are either unsupported by existing data or have yet to be thoroughly tested.”
And I agree that the worst case scenarios are mostly unsupported. Do you disagree with their stance that the assumptions used to hype worse case scenarios “are either unsupported by existing data or have yet to be thoroughly tested”
Please provide the evidence for your disagreement. Otherwise it feels like you are trying to obfuscate the core of their arguments by word-smithing.
Joshua says, “Now I asked you some direct questions about what I bolded. I think those questions make it quite clear what I was emphasizing by having bolded certain points. I notice that in responding you did not answer those questions.”
But I did answer your questions as best as I could. You simply seem incapable of understanding the issues. What you bolded were not questions. So perhaps instead of your excessive use of wordy obtuseness to emphasize “warm” temperatures, please precisely state your question in 20 words or less.
Jim –
I have a few minutes while I’m waiting for someone, so I will add a few things.
One key point in understanding what I was emphasizing is to note the words “primary” and “mortality” in the parts I bolded. There are others as well that you missed in your (wrong) impression of what I was “clearly” emphasizing
Another is to note that the authors I excerpted spoke to
1) the value of popularizing worst case scenarios.
2) the effectiveness of existing communication.
3) recommendation that such communication should be sustained.
4) focusing on existing gaps.
While I don’t doubt that #4 is an important consideration, it seems to me that in your characterization of what they said, you left out, 1, 2, and 3.
A complete and fully accurate characterization of what they said, it seems to me, would have included all those points.
Joshua, I have gone out of my way to try and answer all your questions but you try to accuse me of avoiding your questions which raises my suspicions you are just a “polite troll” desperately trying to punch a hole in the essay no matter how trivial the point. The main point of this essay is coral can rapidly adapt to temperature changes and bleaching is a legacy of that adaptive mechanism. Yet you have yet to comment on the crux of the issue choosing instead to myopically obsess on trivial passages.
So tell me, do you believe the experts that argue coral can rapidly adapt via symbiont shuffling and shifting? The authors of lone sentence that you are stuck on, also wrote,
“Repeated bleaching episodes in the same coral assemblages and the increasing scale and frequency of coral bleaching are cited as evidence that corals have exhausted their capacity to evolve to rising sea surface temperatures (Hoegh-Guldberg 1999). However, few studies have compared the rates of mortality or thermal tolerance within populations in sequential bleaching events. Conceivably, mortality rates may be declining even while the spatial scale and frequency of bleaching is increasing. Indeed, a number of studies suggest that bleaching mortality rates have declined and thermal tolerance has increased in some regions. For example, mortality rates in the Eastern Pacifc were signifcantly lower in 1998 when compared with 1982 and 1983 (Glynn et al. 2001) Similarly, Maynard et al. (2008) found thermal tolerance of three common coral
genera on the Great Barrier Reef to be greater in 2002 than that expected from the relationship between temperature stress and bleaching severity observed in 1998. While there must be upper limits to rates of adaptation, these examples suggest that the evolutionary capacity of corals to respond
to thermal stress is far from exhausted.”
Do you think coral have the capacity to adapt to climate change whether it entails more warming or cooling?
Jim-
==> ” You appear to be obsessing over a single sentence that a case can be made for worst case scenarios.”
Neither am I obsessing, nor am I focusing on a single sentence.
Joshua says, “Neither am I obsessing, nor am I focusing on a single sentence.”
Although that appears to be the case, it is not important. More important is that you are not asking any concise questions nor are you responding to the crux of the essay about the coral’s ability to rapidly adapt via symbiont shuffling and shifting? But perhaps you missed my question so I will repeat.
Do you think coral have the capacity to adapt to climate change whether it entails more warming or cooling?
“…are all known to mediate the bleaching effect and influence the elasticity of the threshold. Therefore, environments with attenuating properties are likely to improve the chances for persistence of corals.”
Many factors are going to go into the bleaching outcomes. Sub-regional variability makes it difficult to boil it down to a one value input. I suppose coral bleaching alarmism is similar to polar bear, bird, butterfly, and penguin alarmism. It is good enough to use one value input? We have another problem perhaps because of the bifurcated outcomes, that is death or survival. Each organism in a large population has their own specific threshold, their own tipping point. Two species of coral in close proximity may exhibit two different thresholds increasing the needed inputs to calculate the expected outcome.
There appears to be a cut and paste error in the paragraph following the heading ‘Multiple causes of bleaching’. The sentence beginning with “In contrast to researchers like Hoegh-Guldberg…” appears again a few lines further down.
Great info. Minor typo: “But or of colder…”
Reblogged this on Climate Collections.
An anecdote:
A friend of mine just returned from Australia where he was a tourist on the Great Barrier Reef. He says he asked the tour guide about coral bleaching and the guide said it was all being overblown by the activists. The friend reported that he did not see any evidence of coral bleaching during his visit.
When some agency says something like it is the hottest month/year on record, all I can do is look at my local condition and say that is not true here. I cannot personally check global conditions, and in reality no one can with any certainty.
So I cannot check global coral bleaching or the success of alternative energy in Portugal.
However when someone reports that something is happening where I live to their foreign audience, I can check.
And that’s the neat thing about the world wide web, it’s world wide and people like me can check “facts” reported about my neighborhood.
Not disagreeing with your primary point but I would note that tour guides would, of course, take their clients to reef areas with robust coral colonies. The opposite goal may be true of some “bleachers”. Cherry-picking of the sea, so to speak.
Reading all the comments..pal review was awesome. Go red team.
Now if Jim D and a couple others stay quiet the echo chamber will be in perfeck harmony.
Steven Mosher at 10:18 pm claims to have read all the comments saying they are all pal reviews and yet at 8:05 pm Joshua questioned the author in a non-pal review way.
Seems like someone needs to read harder.
Steven Mosher: Reading all the comments..pal review was awesome. Go red team.
If there is contrary evidence that has been as well reviewed, please link to it or cite it.
Steven Mosher 10:18 pm
“Reading all the comments..pal review was awesome.”
However Joashua’s 8:05 pm post is not pal review.
Is some harder reading required?
Wow, explicit political acknowledgement. Actual scientists have no politics by design. They don’t eat cookies by design. They have no buddies by design.
Mosher, “Reading all the comments..pal review was awesome. Go red team.”
Well, for a blog post it is pretty comprehensive so judging the review based on the first few comments isn’t very “charitable”.
SM:
You should check your gun sights alignment. For this particular topic, your aim is off a bit.
Or perhaps it was just that hair trigger of yours.
Stephen Mosher,
I certainly can understand how the heat in the kitchen is getting to be a bit too much to bear, what with your fact-free, logic-free and morality-free arguments, which have become the hallmark of the climatariat.
But hey man, if you want to leave, and take “Jim D and others” with you,
http://i.imgur.com/bmwX9UA.png
Jim Steele, thank you for the essay.
I have seen this article twice now and it contains an error. Scott Reef is listed here as GBR (Great Barrier Reef), but Scott Reef is an isolated reef off the NW coast of Australia. It does not affect the assumptions and perhaps provides stronger support. This is because Scott reef lost nearly 80% of it’s coral to a bleaching event but recovered to where it was inside 10 years. This was particularly astonishing because there were no other reefs around to reseed the coral which implies great resilience of corals and really shows that the Green lobby’s claims that the GBR is dying is just not backed up by observations or scientific evidence.
PB, you are right I mistakenly placed Scott Reef in the Great Barrier Reef, sorry. But as you report, it is a great example of coral resilience
Jim
All the talk is always of warm water coral. Here in the UK we also have coral reefs
http://www.marinereservescoalition.org/conservation/uk/uk-marine-life/cold-water-coral-reefs/
They have been growing for 8000 years and apparently are not reliant on sunlight. Our sea temperature varies between around 6 degrees C in a cold winter to around 18C in a warm summer, so quite a temperature range.
Do such reefs suffer due to temperature fluctuation or are they quite different to the reefs you talk about, for example their non reliance on sunlight? In short could they suffer from bleaching and DO they suffer from bleaching?
tonyb
tonyb:
From your link:
No algae, no bleaching.
Hi Climate Reason,
Indeed there are cold water reefs and typically in those regions they do not form a symbiosis with dinoflagellates. As Opluso correctly says “No algae, no bleaching.” Coral are upside down jellyfish that can filter feed and prey on plankton, so I imagine most cold water reefs are determined by plankton availability, but I have not perused the literature on cold water reefs to offer any meaningful insights. In contrast warm water coral live in very nutrient poor waters and must depend on their photosynthesizing symbionts. However during prolonged bleaching events, alternative feeding on plankton can prevent mortality.
Actually, there are a couple of thousand types of “soft” corals and what was formerly known as Gorgonian colonies, do often have symbiotic algae.
It would be interesting to find out how closely related the cold water corals are to the warm water corals. -2C to over 30C is a pretty large range for these endangered critters.
The most important take-away from Jim Steele’s article is that corals display a wide range of ecological adaptations. Not merely between species but within single species and specific local populations. He foreshadowed this with his very first sentence and went on to demonstrate it with an excellent overview of the science.
This provides an argument against climate change being able to extirpate reef-building corals. That is not to say human activities (direct and indirect) are not having a negative impact on corals around the globe. But it is to say that the over-emphasis on gradual GHG warming distorts the picture.
As an aside, I think that the Wikipedia entry on coral bleaching is actually quite good, presenting a balanced view of the issues and the current state of knowledge. https://en.wikipedia.org/wiki/Coral_bleaching
http://www.aims.gov.au/documents/30301/908847/Discovering+Scott+Reef+-+Introduction.pdf/608fe19f-5756-4cd9-82f3-b895a3e82269
Just for fun.
” . . . is updated daily and shows the ocean water temperature as recorded on 25th May 2016
Todays maximum sea temperature: 32.2°C / 90°F (Koh Tao, Thailand) | Todays minimum sea temperature: -1.7°C / 29°F”
Judging from photos, the corals at Koh Tao are alive and well, and the dive shop “Coral Grand Divers” doesn’t seem to be going out of business due to global warming.
On the other hand, the coral Lophelia pertusa prefers temperatures between 6 – 8 C. Corals are found from the tropics to polar regions, and at depths from extremely shallow to thousands of meters.
Alarming statements about coral bleaching need to specify the species of coral (there are at least several hundred species), the definition of bleaching (there are several different conditions referred to as bleaching), and discussion of the various reasons for the various types of bleaching in different species.
A bit of precision might help to avoid some of the argy-bargy on display from time to time.
Cheers.
Corals in Koh Tao are not “alive and well.” This type of ignorant and blatantly incorrect posting exposes the lack of knowledge embodied in the Steele article. I took classes on coral reef ecology in Koh Tao and witnessed wide scale bleaching and coral death in 2014. Here is some background on the damaging impact of coral bleaching in Koh Tao:
http://www.newheavendiveschool.com/articles/impact-el-nio-koh-taos-reefs/
Thailand is currently closing some islands and dive sites due to coral bleaching:
http://phys.org/news/2016-05-thailand-sites-coral-crisis.html
http://www.news8000.com/lifestyle/travel/thailand-dive-sites-shut-over-coral-bleaching/39753444
So, some dive shops and dive sites are “going out of business” despite your assertion.
http://thainews.prd.go.th/CenterWeb/NewsEN/NewsDetail?NT01_NewsID=WNEVN5905220010007
It is frankly shameful that Steele and others here distort the facts to pretend that widespread coral bleaching is either not happening, or not a problem, or can be magically resolved by instantaneous adaptation by symbiotic algae. No wonder Mr. Steele does not publish this specious and misleading propaganda in scientific publications.
It is sad that since global coral bleaching and death is such obvious and clear proof that climate change is happening and is devastating to ecosystems that Steele and others work diligently to distort science to try and pretend that it is not.
Tripp you are looking more like an internet sniper when you say, “Corals in Koh Tao are not “alive and well.” This type of ignorant and blatantly incorrect posting exposes the lack of knowledge embodied in the Steele article.”
Please quote exactly what I said the exposes the lack of knowledge in the essay. If not it is you who is engaging in “ignorant and blatantly incorrect posting”
Well 3 strikes and you are out. Clearly you are just sniping and trolling when you say “It is frankly shameful that Steele and others here distort the facts to pretend that widespread coral bleaching is either not happening, or not a problem, or can be magically resolved by instantaneous adaptation by symbiotic algae. No wonder Mr. Steele does not publish this specious and misleading propaganda in scientific publications.”
Again it is you that continue to distort by creating strawman arguments. No one has ever said bleachng is not happening. Did you even read the essay? Or how bout the title. Again you have yet to quote or refute one of the essay’s distorted facts
I have synthesized what others have published about symbiont shifting and coral recovery. It is not magical and it is not propaganda. It is peer reviewed science whether I or others have published it.
I also followed some of your links. One article talks about dive shops closing due to bleaching and that the government was closing access to the reefs stating,
Bleaching is often temporary and normal. The concern is over mortality and a lack of recovery. So what is the real threat? tourists? Or perhaps rising CO2 cause more tourists?
Coral bleaches, trees lose leaves in autumn, moso bamboo loses leaves in spring. Some “loss” events are annual, others irregular.
In about forty years time all the moso about the world will flower at the exact same time and try to die…
And corals will bleach, sometimes on a big scale. Meant to be. No safe spaces in nature.
mosomoso, Trees naturally shedding their leaves is an excellent analogy to coral bleaching and symbiont shuffling. The more variable the conditions the more likely leaves are to be shed. My favorite plant in the American southwest deserts is the Ocotillo. rainfall is unpredictable where it grows, so depending on the frequency of rain storms, the Ocotillo can shed and regrow its leaves from once 5 or more times in a year.
As cited in the essay, the least amount bleaching has been observed in the warmest waters but where temperatures are less variable. Some of the worst bleaching happens in the eastern Pacific where coral are exposed to both warm El Ninos and cold upwelling, or in more poleward latitdues where winter temperatures can be sometimes be lethal.
In repsonse to warm events, coral can gain acquire warm-tolerant symbionts but shed them like leaves when temperatures cool to acquire new symbionts that can maximize photosynthesis at cooler temperatures. Translocation studies have shown when coral are moved to slightly cooler latitudes, photosynthesis is reduced and unless new symbionts are acquired they die.
More on these ancient organisms, Corals, that date back to the early Triassic:
See, Doubt and Certainty in Climate Science by Alan Longhurst
Thank you very much! I just came up from snorkeling at Tunnels Beach, Kauai and my curiosity was peaked the experience and hearing some talk of global warming impact on the reef which sounded somewhat sketchy. I was thrilled to find your piece. But no discussing with the younger set now, I’m on vacation.
See Bob Tisdale’s essay WUWT May 23rd ‘Say Goodbye to the 2015/16 El Niño’. There is an animated GIF showing daily global SST change (as reported) from March 26 to May 24th or so this year.
The El Nino region of the tropical Pacific shows changes that have been attributed by many to involve redistributions of hot and cold volumes of near-surface water. From other data, the redistribution is slow, going from one extreme to another over months rather than days.
I looked for a long time at the patterns in the Mediterranean Sea. There is less scope for large redistributions in its relative confinement. SST patterns change over several days rather than several months. Is this fast change a consequence of global warming? I suspect not.
If not, then what?
One has to come back to cloud formation as a likely contender. It is intuitively and physically likely that cloud cover can play with these SSTs in the Med Sea.
If clouds can do it there, clouds can do it where there are coral reefs. So, in discussions of bleaching mechanisms, I’d tend to downplay El Nino cycles and concentrate more on short term changes. Surely there is data from places on the Great Barrier Reef showing daily insolation ( as opposed to temperature maxima, for example).
Can anyone help with references about cloud weather effects on reef properties like bleaching?
Geoff, you are correct that clouds also play a major part in bleaching and I only mentioned it in passing that “high irradiance from clear skies ” also causes bleaching. I do not know of a paper off hand that focuses on clouds but heavy cloud cover is often mentioned in several papers as the reason why some reefs were NOT bleached by the severe El Ninos
Here is a link to a paper that mentions incidents of bleaching or lack of bleaching attributed to clouds
http://gefcoral.org/Portals/25/BWG/EcologicalMonograph2007.pdf
example
“reefs of Mauritius have some of the highest SSTs in the southern tropical Indian Ocean (1.70–1.82) and were among reefs least affected by the 1998 bleaching (McClanahan et al. 2005b). The temperature history of the sites has been overlooked and hardly reported, and the low bleaching in 1998 has been attributed to the cooling effect of Cyclone Anecelle, high
cloud cover, and rainfall”
Thank you Jim. Yours is a neat essay.
Geoff.
Jim,
Great article, but:
“Perhaps CO2 concentrations are higher now than over the last 300 Ma.”
On the contrary, CO2 levels have been higher than at present for most of the past 300 Ma. See Figure “Phanerozoic Carbon Dioxide” here:
https://en.wikipedia.org/wiki/Carbon_dioxide_in_Earth%27s_atmosphere#Past_concentration
Gareth, I agree that CO2 was most likely higher throughout the past 300 Ma, but there is considerable debate and limited proxy data. I said “perhaps” only to accept the possibility, but even if it were true, it suggests low climate sensitivity. Models driven by CO2 trying to explain the warmth of the Mesozoic require unsupported high concentrations or unsupported high climate sensitivity. Claimed CO2 effects are rife with contradictions,
Well, any time historic temperatures have been cold, there has been a large land mass over the south pole.
Unless CO2 moves land masses it doesn’t have a lot to do with historic temperature.
PA, I agree that the position of the continents were the primary driving force in the earth’s cooling. As I wrote in the essay the “Antarctic Refrigeration Effect”, when the continents separated from Antarctica and the Antarctic Circumpolar Current inhibited poleward movement of warm waters that’s when Antarctica began to cool and the extruded brine from sea ice formation began cooling the ocean depths.
http://landscapesandcycles.net/antarctic-refrigeration-effect.html
Pingback: Coral bleaching debate — Jim Steele | Climate Etc. (Judith Curry) | Taking Sides
The Great Barrier Reef Wars are one of the dozen or so “scientific controversies” of our day. [ The Climate Wars are, of course, another — The Salt Wars, The Obesity Wars, etc. ]
Steele is correct in his data….and the fact that coral reefs (in general) and the Great Barrier Reef, in particular, and the science surrounding them is being used as a political football and the facts grossly misrepresented to the public for various purposes that I do not understand — but at least efforts to increase the “importance and urgency” of their special field — and this ensure continued research funding.
The GBR Wars have been ongoing for many many years and are a continuing feature of Australian environmental politics. The GBR Wars have slopped over to the rest of the world and are being used to whip up associated furor over Global Warming, Plastics in the Sea, coastal pollution.
The pragmatic problems for reefs are over-fishing and illegal fishing methods (drag nets over reefs, dynamite, cyanide), real pollution from shore for near-shore reefs (sewage, storm drains, industrial and agricultural chemicals), and natural occurrences: cyclones, diseases, COTS, El Niños, cold snaps, etc. Most destructive over-fishing is a by-product of local poverty and lack of alternative employment.
My personal experience is in the Caribbean, where near-shore reefs have been badly damaged by pollution from shore and (centuries of) over-fishing. Reefs away from concentrated human habitation (cities) are doing better and where fishing is prohibited or limited, such as in the Exuma Cays Land and Sea Park (http://exumapark.org/), the reefs have rebounded magnificently.
Great Barrier Reef omitted from UN report amid claims department intervened
Australia is not mentioned in a major UN report about the impacts of climate change on heritage sites, despite being home to the world’s largest reef.
http://www.sbs.com.au/news/article/2016/05/27/great-barrier-reef-omitted-un-report-amid-claims-department-intervened
Perhaps this article has had an impact? ;-)
The problem was miraculously solved by the Australian Government.
“The Turnbull government has forcibly removed any reference to Australia from a new UN report – In a statement to news.com.au, the Environment Department confirmed it asked for references to Australia to be removed, saying it would have a negative impact on tourism”
http://www.news.com.au/technology/environment/climate-change/the-turnbull-government-has-forcibly-removed-any-reference-to-australia-from-a-new-un-report/news-story/30425c0cf0e19987e2fa8ba22731dc5d
See how easy it is to solve the problem of Climate Change. I just can’t wait till Trump shuts down NASA, NSF, NOAA and the EPA. I think he should remove those annoying cancer warning labels on cigarettes too. And come on, do we really need seat belts? It will be just like 1956 again.
jacksmith4tx,
Maybe you should send your thoughts to Trump. I’m not sure whether he would agree with your proposals on cigarettes or seat belts, but I suppose the majority is supposed to rule. It’s worth a try, I suppose.
Or just get the Australian government to tell the UN what to do. It seems that Australia runs the UN’s climate change agenda to a greater or lesser degree, doesn’t it?
Why waste money on the UN? Just seek advice from Australia!
Cheers.
I share your concern in the sense that I never approve of governments’ censoring information. On the other hand it is just as easy for government entities to create false climate alarm, so I am curious how much of their reaction remove Australia’s input was evoked by some researchers over dramatization and false characterization of recent bleaching as gloom and doom.
In the USA Clinton and NASA hyped Parmesan’s butterfly study to suggest climate change was killing butterflies at low elevations and southerly latitudes. All the other experts reported it was urban sprawl’s removal of habitat that was the issue and later the colonies Parmesan reported as extinct have returned. Yet Clinton brought her to the White House http://landscapesandcycles.net/climate-doom–parmesan-s-butterfly-effect.html
The point is governments can both create or shut down climate change paranoia. Our job is to critically analyze the propagnada.
See how easy it is to solve the problem of Climate Change. I just can’t wait till Trump shuts down NASA, NSF, NOAA and the EPA. I think he should remove those annoying cancer warning labels on cigarettes too.
Given that climate change is a political problem not an actual problem that would fix it. And the government funds saved could reduce the debt or go to funding honest science instead of “global warming” science.
I want to apologize for my snarky comments about Trump. I really have no idea of what he will do once he is president (and nobody else does either). What ever he decides to do it will be pretty damn hard to stop him. I don’t really think we will need a congress or supreme court after we elect Trump, they will just be in his way. Most americans hate congress anyway so who’s going to stop him?
brandonrgates,
On the one hand, you write –
“I’m not a coral expert. Not even close.”
On the other hand, you seem to be pretending to some expertise in the field, otherwise it would appear more logical to ask questions in order to increase your knowledge, wouldn’t you agree?
The Warmist mantra of “energy production by burning stuff must be evil” is losing its impact. Predictions of imminent doom from increased temperatures due to human activity in various areas seem to be overstated.
It seems you are not only not a coral expert, but not an physics expert, either. Maybe you might care to state your claims to expertise in other areas. Have you any?
If you seriously desire knowledge in an area where you think I might possess more than you, feel free to ask. I won’t just demand that you look it up on Google. I assume you would do that before you asked me for assistance. I’m always glad to help a serious seeker after knowledge.
Cheers.
Here is the science
Given corals’ evolutionary history
Given coral survived warmer oceans during the Medieval Warm Period
Given the there is a vast community of symbionts including clade D with many species adapted to warmer temperatures
Given that coral can shuffle and shift their symbionts and acquire warm adapted symbionts
What is the evidence that coral can not adapt to a rise in temperatures,such as the 2.1C rise during the Holocene Optimum???
The global warming alarmists have no argument and no evidence, other than their paranoia. So all they can do is dishonestly attack the science presented in the experts’ adaptive Bleaching Hypothesis and make snark comments.
The only honest statement from Brandon is ““I’m not a coral expert. Not even close.”
Jim Steele,
Would that you would be even that forthcoming.
Brandon may indeed say, “I’m not a coral expert. Not even close.”
But then, when asked to marshall evidence in support of his claims, he bemoans, “It’s my job to teach this entire forum about corals. One thing I do know for sure, y’all aren’t paying me enough to do your homework for you.”
https://judithcurry.com/2016/05/24/coral-bleaching-debate/#comment-786285
This sort of passive-aggressive behavior is quite revealing.
Here in Mexico they have a saying for it: Hay gente que es purita mula y se cree caballo de carrera. (There are people who are purebred mules, but believe they are race horses.)
Glenn Stehle,
One might even argue that it’s indeed appropriate for me to openly volunteer that I’m not a coral expert. That lets readers know that I’m not attempting to trade on my personal expertise, and to be appropriately skeptical of my own interpretations.
No other evidence is required to point out that Steele misrepresented what his literature citations said. Nor that Steele’s overly-confident predictions that corals will adapt come what may does not follow from his manufactured premises.
Surely you’re not suggesting that it is indeed my responsibility to any homework for you.
I am not a coral, Glenn. Some, certainly not me, might say that not playing the ball reveals the vacuity of your non-argument. Same could be said of Steele’s frequent and ongoing well-poisoning.
However, do please carry on. Especially continue vainly contradicting the fact that I did indeed provide additional citations even as I wrote that it wasn’t my duty or requirement to do so. I’m always thankful for an opponent who tells easily falsifiable porkie-pies.
Brandon lies again, saying, “No other evidence is required to point out that Steele misrepresented what his literature citations said. ”
More precisely there is .no evidence that I misrepresented the literature. Brandon is a well known internet sniper whose mission has always been to try assassinate the character of all skeptics. Brandon Gates is has been shown time and time again that he will lie and fabricate to do so!
Jim, you repeatedly refer to corals adapting to changing temperatures over thousands and millions of years. However, the man-caused rise in temperatures in this century is faster than average and caused by man. And it will get worse if we don’t reduce greenhouse gas emissions:
http://earthobservatory.nasa.gov/Features/GlobalWarming/page3.php
“As the Earth moved out of ice ages over the past million years, the global temperature rose a total of 4 to 7 degrees Celsius over about 5,000 years. In the past century alone, the temperature has climbed 0.7 degrees Celsius, roughly ten times faster than the average rate of ice-age-recovery warming.”
Your immediate adaption theory is hokem and corals cannot adapt to the rapid temperature rises caused man’s greenhouse gas emissions.
Tripp again obfuscates saying
The focus of the essay has been on how rapidly coral can adapt. Arguments that coral bleaching is evidence that coral have not bleached is pure rubbish and gibberish. Cloud cover can cause one reef to bleach but not its neighbor. Later the reef that never bleached and thus never underwent symbiont shifting then bleaches. Observations have also shown some species on a reef bleach heavily during one episode and then become resilient during the next episode, but then a different species bleaches.
Tripp blathers
It is not ‘my” theory but a theory prposed by coral experts since 1993 and has been gaining increasing support.
Tripp I must say you persist in denying observations and evidence to simply fit your theory and then to insult people who provide the evidence to show how wrong you are.
If you are so sure Tripp of your catastrophic beliefs then take my bet
Oops too late at night. The sentence above “arguments that coral bleaching is evidence that coral have not bleached is pure rubbish and gibberish” should have “arguments that coral bleaching is evidence that coral have not adapted” is pure rubbish
Ragnaar, Your linked article hints at exactly what the adaptive bleaching hypothesis suggests. Abrupt warming by an El Nino bleaches the vulnerable polyps. The polyp symbiosis that were adapted survived and will restore the reef within a decade. The articles says “There were exceptions, though — tiny pieces of baby coral that are beating the odds. And those exceptions could give scientists insight into how to make sure climate change doesn’t completely destroy reefs around the world.”
They call incorrectly cryptic polyps “baby coral” but the idea is the same and we can expect similar recovery as those I cited”
“At Northwest Australia’s Scott Reef, the upper 3 meters lost 80 to 90% of its living coral and the disappearance of half of the coral genera. Yet researchers observed, “within 12 years coral cover, recruitment, generic diversity, and community structure were again similar to the pre-bleaching years.” A similar long-term study in the Maldives observed a dramatic loss of coral during the 1998 El Nino but by 2013 the reefs also had returned to “pre-bleaching values”. Although a reef’s recovery sometime requires re-colonization by larvae from other reefs, a process known as re-sheeting or Phoenix effect can facilitate a reef’s speedy recovery. Often a small percentage of living “cryptic” polyps with a more resilient symbiotic partnership were embedded within a “dead” colony and survive extreme bleaching. They then multiply and rapidly “re-sheet” the colony’s skeletal remains.”
Pingback: Jim Steele ja korallien haalistuminen | Roskasaitti
It is easy to recognize insincere internet snipers based on their self contradictions and schizophrenic cherry picking of quotes out of context.
Our current troll has repeatedly attacked my comment that because coral survived temperature 2.1C degrees warmers than today during the Holocene Optimum and the observations of their resilience and rapid recovery, that it virtually guarantees they will survive a similar warming. The is based on multiple lines of past and present evidence.
In contrast, the trolls accept unsupported fear mongering statements by Hoegh-Guldberg that “as much as 95% [of the world’s coral] may be in danger of being lost by mid-century.” That fearful speculation is based solely on models that failed to simulate Holocene temperatures and account for the Holocene conundrum, the persistent denial of the emerging evidence that coral can rapidly evolve, and dismissal of observations that many bleached reefs have fully recovered in less than a decade.
Another example of the internet snipers obfuscation conflates separate issues into muddled confusion and misdirection.
For example, “Your “virtual guarantee” of coral survivability ignores other causes which you yourself say are responsible for *most* coral mortality. Your temperature-centric guarantee doesn’t follow from its own premises, Jim. That’s not my issue, it’s yours.”
The essay is about adaptive bleaching and symbiont shuffling and shifting that creates greater resilience.Again it is the alarmists who argue that bleaching is the result of of coral at the limit of their thermal tolerance. The trolls accept alarmists thermal myopia but suggest any rebuttal to that limited view of adaptation does not address the other issues. Such misdirection actually reveals their total foolishness
Adaptive bleaching and symbiont shuffling and shifting can never make coral more resilient to hurricanes or predators or to dynamite fishing. Only a very stupid troll or a very determined internet sniper would ever try to conflate those issues. The mechanism that has allowed coral to rapidly recover from destruction from those other factors like cyclone damage is resheeting. That and that is the other mechanism that has allowed coral to recover through the millennia.
My guess is trolls like Brandon Gates blindly believe Hoegh-Guldberg’s prediction that 95% of the coral will be gone in 34 years. It wont be long before nature will expose who is misrepresenting coral biology.
The Baker 2003 excerpt confirms everything presented in the essay. It acknowledges many unanswered questions, yet since 2003 the emerging evidence continues to support the adaptive bleaching hypothesis and answer some of those questions as confirmed bythe links to papers published a decade later.
Is it “possible” that coral may not be as resilient as the emeging vidence suggests. Of course! That is always the case. But what seems less and less likely is Hoegh-Guldberg’s fearful claims that we will lose 95% of our coral by mid century, a claim that Gates seems to embrace andhas yet to question. Gates only challenges, ad nauseum, any science that suggests resilience and optimism. But that’s what trolls do. Gates should heed Hutton’s advice.
What Steele says:
What the paper he cites says:
Despite the expressed difficulty of making meaningful conclusions as stated in his own citation, Steele concludes in comments:
His conclusions don’t follow from his premises, especially since his premises are based on misrepresentations of citations he himself has provided.
What Steele says:
What one of the citations he references, Baker (2003), says:
Despite all the unanswered questions in the citation he provides above, Steele (over)confidently concludes in comments:
ROTFLMO Brandon Gates’ desperate attempt to regain a smattering of credibility and continue his attack on my credibility, is typical of internet snipers from Slandering Sou’s school of skeptic bashing, and again reveals their/his total scientific ignorance.
Agreed I did not discuss every issue presented in the paper to which I (and Gates) linked to, “Coral—the world’s most diverse symbiotic ecosystem”. But bleaching is caused by the loss of eukaryotic photosynthesizing dinoflagellates from the genus Symbiodinium. My link to that paper was merely to confirm that the number of identified symbiodinium species has increased from one to hundreds as expected by the Adaptive Bleaching Hypothesis.
The paper’s most relevant quote is
As typical for all peer reviewed papers, scientists cite a paper that confirms their assertions. Those citations are never meant to be a synthesis of the entire paper being cited. Brandon Gates is not only NOT a coral expert but clearly he is not a scientist or he would realize this. But that is not Gates’ major gaffe.
The cited paper is a review of all the organisms that have a symbiotic partnership with coral. In the context of bleaching, prokaryotes (i.e. bacteria, and archea), as well as viruses play a role in chemical recycling but their roles were irrelevant to the bleaching issue being discussed. But Gates in his desperate attempt to attack my character, grabbed at the first sentence that he could no matter how irrelevant to attack my confidence in corals’ survival quoting
But Gates clearly doesnt know the difference between prokaryotes and eukaryotes. His quote had nothing to do with the symbiodinium species being discussed and the difficulty identifying prokaryote diversity is well known by every researcher, but to the likes of Gates
I must admit there is a degree of pleasure when hateful ignorant trolls hang themselves and reveal their total ignorance that highlights their dishonest tactics. ROTFLMAO!!!
Jim Steele,
Yes, leaving things out is the pattern I’m pointing to.
And my link to that paper was merely to confirm that you again glossed over some salient details.
As I’ve already volunteered, I’m not a coral expert by any means. Nor am I a working scientist. But I do know the difference between refereed primary literature and this essay of ours. If we must make this about personal expertise, I don’t find on your bio any mention of any working experience with corals. Perhaps it needs an update?
Your bio also mentions that you’ve taught science at the high school and first- and second-year university level. I sincerely hope your method of teaching didn’t include ROTFLMAOing at your students for not being scientists or bird experts. Just sayin’.
You’re funny. I’d explain the difference, but then you’d claim I simply Googled it, so I see little point. Besides, the distinction not relevant to the argument I was making.
Yes, you are correct. However, I didn’t say the prokaryotic species had anything to do with Symbiodinium now did I. More misrepresentations from you, tsk. Let’s read the concluding paragraph of Blackall et al. (2015):
Wording of the first sentence suggests that prokaryotic holobionts generally outnumber eukaryotic and viral holobionts. The naive expectation is therefore that the potential role of prokaryotic species shouldn’t be neglected just because the topic of a particular essay is on the symbiotic relationship corals have with various eukaryotic dinoflagellates (e.g. Symbiodinium). And indeed, the body of the paper suggests that knowledge of the role of prokaryotic holobionts is still lacking:
My critique of your essay remains that it is far from a *comprehensive* analysis, thus rendering your overconfident virtual guarantees dubious, if not wildly wrong.
Jim Steele,
I’m glad you brought that up. In your essay you write:
One of your citations, Baker (2003) contains no fewer than nine references to papers including Hoegh-Guldberg on the list of authors, including the “infamous” 1999 paper:
So according to your argument above, Baker is one researcher who cites Hoegh-Guldberg in support of his own assertions. However, it’s not that simple, there is a tenth citation Baker (2003) makes, Baker AC. 2002. Is bleaching really adaptive?—reply to Hoegh-Guldberg et al. Nature 415:602 This suggests that Baker and don’t always see eye-to-eye (domain experts often don’t, robust scientific debate relies on this fact). I chased down the reference and could only get the abstract:
Some major disagreement, that one. A worthy exercise is to read through Baker (2003) and note when Hoegh-Guldberg is cited in support or contrast. My own reading suggest that Baker doesn’t think Hoegh-Guldberg is any kind of wild-eyed irrational catastrophist, but rather a credible and trusted colleague who happens to have some differing ideas which they sometimes debate in peer-reviewed primary literature. Which is my lay understanding of how science is exactly supposed to work.
Let’s not forget Baker’s conclusions in the 2003 paper:
Stands in pretty stark contrast to your virtual guarantees of coral survivability come Hell and/or high water.
It’s unlikely that massive coral bleaching would be caused by steady temperature rises of even a degree or so. Temperature-caused bleaching would be scattered, at least at first, as individual corals have differing top-hat susceptibility. This is not observed but mass region deaths are, extending from the northerly regions (the most affected). It’s like a poisonous current was flowing southwards …. and that is the case.
The poison is a mixture of H2S and sulphuric acid (which is far more acidic than CO2/H2O) resulting from frequent volcanic activity at PNG and Vanuatu at that time, 2011 to 2015. Anti-clockwise currents from those regions bring the poison – and warm water, thence rain – down along the Qld coast, taking about 2 weeks.
[H2S is discharged from undersea volcanoes – in this case, along PNG north coast and from both sides of Vanuatu – and is about as toxic as cyanide to all oxygen breathing organisms. Both chemicals bind to the Fe in haemoglobin.]
(Australia’s so-called “El Ninos” also arise here – which cause coral bleching. There’s a bit of preliminary in “Planet Earth Climate Topics at my website pjcarson2015.wordpress.com)
Here is what Steele says:
Here is what Hoegh-Guldberg (1999) actually says:
That’s ten not-so-simplistic conclusions, not three simplistic assumptions. Warming is listed as the primary driver of increased bleaching, not the only one (falling ocean pH and SLR are also mentioned). Bleaching *is* expected to increase due to global warming (Steele isn’t *always* wrong). Finally, these two statements warrant a closer comparison:
Steele: c) coral cannot adapt quickly enough to temperatures projected by climate models.
Hoegh-Guldberg: 4. Corals do not appear to be showing any signs that they are able to acclimate or adapt fast enough to keep pace with these changes in water temperature.
There’s a big difference between saying something is not showing signs of being able to adapt, and that something cannot adapt quickly enough. Here’s what over-confident and overly-simplistic hype actually looks like, courtesy of Steele in comments above:
Brandon dishonesty continues as heis working awfully hard trying to discredit the essay but excerpts from every link only support all the essay’s points.
Brandon simply numbers a hodge podge of evidence, speculation and empty assertions and alternative wording to suggest “The bases for his claims relied on 3 simplistic assumptions ” was wrong. That there really 10. ROTFLMAO Gates is just a dishonest word-smithing troll who is clueless about the biology.
Steele writes:
Here is what one of the supporting citations, actually says:
Hoegh-Guldberg (1999) is indeed questioned throughout the body of the paper. I’m not finding Hoegh-Guldberg being singled out for accusations of “popularizing worst case scenarios”, however.
As for “persistently misunderstanding and misrepresenting the adaptive bleaching hypothesis”, Maynard et al. have this to say about ABH:
Compare Steele’s statements from comments above:
Sure looks like premature heralding symbiont shuffling as a panacea for reefs threatened by climate change to me. It continues looking like many of Steele’s citations in his essay either don’t say what he says they say, or directly contradict his simplistic, overconfident and ill-formed conclusions.
Pingback: El coral sufre por el “cambio climático” … ¿de verdad? | PlazaMoyua.com
“A bittersweet victory for an El Niño chaser” by Kim Cobb (Georgia Institute of Technology) at NOAA, 27 May 2016. Excerpt:
It has been a tough year for the world’s corals, as warm ocean temperatures associated with one of the largest El Niño events on record drove extensive coral bleaching and coral mortality. As someone who uses coral skeletons to reconstruct El Niño events in the distant past, it was a decidedly mixed year, filled with the exhilaration of a once-in-a-lifetime scientific opportunity as well as profound grief for a reef I’ve been diving on for 18 years. Let me explain.
… We arrived to witness the wholesale destruction of the reef that I know like the back of my hand as my primary research site for the last 18yrs. While a few corals were recently bleached, the vast majority of the corals had succumbed months ago, already coated in a thick layer of red-brown algae. They will never recover, and will be reduced to white coral sands in the coming months. You can read more about my personal experience of this mass mortality event here and here.
It was particularly hard to see 50-60yr-old coral colonies completely dead – coral colonies that I had drilled for the Showtime documentary on climate change called “Years of Living Dangerously”. Our cores indicate that some of these old colonies had bleached during the 1997/98 El Niño event, but none had died off completely. At face value, therefore, it appears that the 2015/2016 event was far more devastating to the Kiritimati reefs than anything in the recent 50-60 years.
Our future work will help to understand if the magnitude of the 2015/2016 El Niño event is exceptional not only in the context of the 20th century, but over the last millennia or more. Such research is critical to the community’s quest to identify if, and how, ENSO is responding to human-caused climate change. The implications for society are significant, for as these last few months have demonstrated, corals aren’t the only living organisms that are sensitive to ENSO extremes.
Will Brandon Gates accept a $500 bet
Hoegh-Guldberg made the “simplistic, overconfident and ill-formed conclusion” that. “as much as 95% [of the world’s coral] may be in danger of being lost by mid-century.”
Based on evolutionary history and the emerging evidence that I presented in the essay that has yet to be refuted by the troll, I shared my optimism that “There is a pool of “pre-adapted” symbionts that can be shuffled and shifted, their adaptive mechanisms virtually guarantee that coral will be able to adapt to the same higher temperatures they survived during the Holocene Optimum.”
So I will bet Gates that by mid-century 2050, Hoegh-Guldberg’s fearful ill-informed conclusion will be proven very, very wrong.
Jim Steele,
Here’s what Baker (2003) actually says:
Here is what Maynard et al. (2008) actually says:
In contradiction to your own citations and in defiance of all that is honest and rational, you continue to issue virtual guarantees:
I have refuted you several times by reading your citations and reporting what they actually say. If that’s trolling in your book, I’ll be a coral’s uncle.
Why is this troll so persistent with his disinformation?
Gates claims
ROTFLMAO Truly a legend in his own mind who has been show to be wrong and doshonest throughout this comment section. Gates simply embraces the propgandist’s tactic that if you tell a lie often enough people will believe it.
All Gates’ quotes about uncertainty only reveals that Hoegh-Guldberg’s simplistic ill-formed fear mongering that 95% of the coral will be gone by mid century is truly bogus. And that’s why Gates refuses to bet . But Gates is quick to embrace all the gloom and doom so the troll desperately cuts and paste irrelevant excerpts to suggest only confidence in the more optimistic emerging science is misplaced.
Again Gates’ excerpt from my citation summarizes the essay’s main point
Of course it is not a panacea for dynamite fishing and polluion. That’s never been the argument. Just another Brandon Gates’ dishonest misdirection.
Jim Steele,
Keep telling us that and even you may begin to believe that you didn’t just write:
One good reason to not take your 35-year bet comes courtesy of Hoegh-Guldberg et al. (2008). Clearly not as … optimistic … as you are, but not so doom-and-gloom as your well-poisoning against him suggests. But hey, I’m no coral expert … I’m not even a scientist … so what do I know.
Apparently my biggest sin here has been to focus on your arguments.
Brandon Gates replies with more nonsense
I totally stand behind my statement and you have still not come close to refuting. Every excerpt from my citations support my claim. Other researchers like Smith echo my expectations with very similar wording.
Gates is clueless about the difference between refutation and uncertainty. Only a fool or malevolent troll would foolishly and repeatedly portray one scientist’s uncertainty as a refutation of the evidence that evokes confidence in another.
brandonrgates | May 29, 2016 at 5:44 pm |
Apparently my biggest sin here has been to focus on your arguments.
So far all you have done is bring spitballs to a gun fight.
You haven’t seriously addressed any his points and mostly blow smoke. I haven’t seen you present any hard facts, but must admit I really haven’t been paying attention.
As far DNA is concerned, as little as scientists know about climate.they know even less about DNA.
It is estimated that 3% of the human genome does something we recognize as useful but up to 80% is active (has a purpose).
Since we don’t know what the majority of DNA does, claiming we can predict the adaption response of organisms is sort of futile.
Further there are hundreds if not thousands of Zooxanthellae species (wiki says the true number may never be known). With that much choice corals could adapt to almost anything.
Wrong!
Although there’s certainly a great deal left to be learned about both.
Function, not “purpose”. Only IDi0ts talk about “purpose” WRT evolutionary theory.
And there’s a lot more than 3% of DNA for which some speculative function can probably be assigned, even if it’s just providing spacers between transcription factor binding sites.
Especially when it comes to interactions among symbiotic partners.
But IMO everybody’s arguing past each other here. There seems to be a general agreement that there’s a good chance that the coral species involved in the reef ecology can adapt, but no certainty.
Either way, that says nothing about whether the complex, presumably climax, ecosystem can adapt. Even if it does, it might do so with a different population (“species” or sub-species) of coral in any major niche.
And another question: what if it doesn’t? What if major equatorial coral reefs essentially die? So what? Best evidence is that the general ecosystems will probably move polewards, so it isn’t as though the phenomenon itself will be lost.
Either new beds of algae will colonize the dead reefs, probably resulting in the emergence of brand new complex ecosystems, or the reefs will dissolve, which will aid in buffering atmospheric CO2. Either would be a net benefit, especially when the current ecosystems will probably survive in new locations.
AK,
IMO, Steele isn’t arguing so much as he’s deflecting my observations that he’s overstating the literature support for the Adaptive Bleaching Hypothesis, especially with respect to warming. IOW, he’s being overcertain, so I don’t see that he’s in general agreement with your statement.
One thing that may have gone missed in the fracas here is that, let’s say, Steele doesn’t have much … confidence … that warming is a function of rising CO2 levels because … models. In essence, his argument here is *even if* there continues to be some *near-term* warming, corals will be able to hack it. Unless I missed something, he’s been conspicuously silent on the implications of long-term warming, I *presume* because he doesn’t believe long-term warming is in cards *even if* nothing is done to reduce CO2 emissions to near zero over the next half-century.
For the record, I think the ABH is well-argued in literature so far as my lay eyes can tell, but when even its supporters conclude that it’s not a panacea which virtually guarantees corals will adapt come what may, I tend to listen. I synthesize those qualifiers about ABH with prior research showing that corals dissappeared from the fossil record in all the Big Five extinction events. Obviously that we have corals today means that they didn’t all go extinct, but the fossil record suggests that they have been … virtually … wiped out in past periods of rapid environmental/ecological change.
In sum, corals are tough buggers to all but eradicate, but not impossible. That they are already dying due to numerous factors tied to human activity suggests a call for concern and *very likely* modification of our behavior so as to preserve them. That includes many various other activities not strictly limited to our present rate of CO2 emissions.
Wait a minute! I thought you were
the oneinsisting on considering the relative speed of warming. As long as it doesn’t go too fast, doesn’t it appear that coral can adapt?Wrong! IIRC there’s good evidence that “stony coral” actually re-evolved (from generalized several times after mass extinction events.
It may be that modern (post-Triassic) corals are descended from earlier calcifying corals, but given the complexity of coral evolution it can’t be considered certain.
In addition, we should remember that “today’s situation, where scleractinian corals dominate, is more of an exception than a rule.”
http://www.ucmp.berkeley.edu/porifera/images/poriferafr.gif
AK,
Among other things like *rate* of SLR, reduction in ocean pH, polluting runoff from land, overfishing, etc. It’s not just about absolute temperature or rate of temperature change that factors, and this post is not the first time I’ve pointed that out on this thread.
You tell me. Both of these references are supportive of the ABH:
Baker (2003):
Maynard et al. (2008):
Neither of those references suggest to me that ABH remotely guarantees that corals will handle *expected* rates of warming. But sure, given a zillion years to adapt to a change in temperature, they might very well be able to go above previously unseen absolute temperature thresholds. Somewhere in the middle of little to no change and instantaneous change is a rate that most corals would tolerate. I’m not presently convinced that anyone knows what that happy medium is, if it’s indeed even knowable.
Ok, fine. One point I was implicitly making is that just because a certain class of organisms disappears from the fossil record does not necessarily mean they went fully extinct. Still, I don’t understand how your point here does anything to address my explicit argument that corals have been known to be all but wiped out by sufficiently rapid/radical environmental changes. Hence, ABH is nowhere close to being a “virtual guarantee” that corals will be able to handle Hell or high water because … shifting/shuffling adaptations.
Just to be abundantly clear, that doesn’t mean I think ABH is bunk. Only that it’s not a panacea as Maynard et al. put it. Certainly not a “virtual guarantee”.
AK | May 29, 2016 at 6:58 pm |
As far DNA is concerned, as little as scientists know about climate.they know even less about DNA.
Wrong!
Only IDi0ts talk about “purpose” WRT evolutionary theory.
Well, at least you didn’t argue there is just one species of Zooxanthellae.
http://psychology.wikia.com/wiki/Noncoding_DNA
For example, in the genomes of humans and mice, which diverged from a common ancestor 65–75 million years ago, protein-coding DNA sequences account for only about 20% of conserved DNA, with the remaining majority of conserved DNA is represented in noncoding regions.[18] Linkage mapping often identifies chromosomal regions associated with a disease with no evidence of functional coding variants of genes within the region, suggesting that disease-causing genetic variants lie in the noncoding DNA
80% of the conserved DNA is in non-coding regions.
“Suggests that disease causing…” Suggests??? Really??? That sure sounds like they are certain what is going on.
There is a good deal of “non-coding” DNA that some function has been found for. There is also a good deal where they’re still looking.
I never said “they are certain what is going on.” Since you obviously didn’t read the link I gave, I’m not going to spend time digging up links to prove it, but the most commonly accepted theory today is that it’s the control regions that are involved in most mutations involved in disease and typical (adaptationist) evolution.
Obviously, if control regions (which are not coding regions) are the primary mechanism of evolution and genetic disease, they are going to be conserved in much the same way as coding regions.
And that doesn’t even start to get into the other types of functional non-translating RNA that has been discovered over the last decade or two. All of which is more-or-less conserved.
Ak says
AK of course there is never any certainty whether rising CO2 will push temperatures to extremes or if coral’s adaptive mechanism can handle climate change whether or not it is natural or not. However based on their evolutionary history and the emerging evidence from symbiont shuffling and shifting that I presented in the essay I have come to believe that coral survival throughout this century is a virtual guarantee. However that certainty was never the point of the essay. The essay simply provides details of the emerging science about coral adaptive mechanisms. Only time will tell if my confidence is misplaced. And only a troll would harp on that one comment ad nauseum as if it was proof of anything.
Agreed my virtual guarantee stands in stark contrast to media hype that states with certainty that coral will go extinct by 2100 or Hoegh-Guldberg’s 2014 statement that “More recently, global impacts from ocean warming and acidification have become increasingly apparent, with very dire consequences if our current emission trajectory continues unabated. Looking across the world’s coral reefs, 75% are threatened and as much as 95% may be in danger of being lost by mid-century” (In fact the most recent research suggests there has been no impact from lower pH and any effect is likely insignificant, but that is another debate discussed in the next essay)
Could my virtual guarantee be proven wrong? Of course. Nothing is 100% Likewise Hoegh-Guldberg’s and all the global warming extinction claims can be wrong. Putting aside the red herring issue of “misplaced certainty”, the question we must ask is, “does the case for resilience or the case for extinction have more support? Hoegh-Guldberg has dismissed all the emerging evidence to argue symbiont shifting will not allow coral to raise their their thermal tolerance limits. That stands in stark contrast to evidence reported in this essay that has synthesized the growing evidence in the peer reviewed literature that symbiont shifting is real and increased resilence is being observed. But this essay puts me in the cross hairs of skeptic bashers like Slandering Sou/Brandon Gates. It is easy to see the confidence issue is a red herring because Gates never criticizes Hoegh-Guldberg’s for over confidence that 95% of the coral will be lost by 2050, because Gates embraces gloom and doom certainty.
You suggest we are talking past each other, but that has been Gates’ intention all along. That has been a Slandering Sou and Gates standard tactic for as long as I can remember. An essay designed for a web post can never cover all the details about coral, or any other topic. So malevolent internet snipers can easily fabricate a trumped up charge of wrong doing and misrepresentation by arguing an essay did not address all the information. The only counter easure would be to write an encyclopedic tome. It would take many many books to cover everything about coral, so creating their strawman, is a tactic easily wielded by any internet sniper.
HIs intentions are readily discerned as Gates first entered the discussion by dishonestly accusing me of misrepresenting a citation by Kleypas. I simply said the least amount of bleaching had occurred in the warmest waters and that is exactly the statement made by Kleypas. But Gates purposefully tries to talk past that point by suggesting I didn’t discuss all of Kleypas’s paper and the ocean thermostat hypothesis. Indeed Kleypas suggest the ocean thermostat hypothesis is why the warmest waters exhibit the least amount of warming. It supplies more evidence of why Hoegh-Guldberg’s 95% fear mongering is very unlikely and I would have loved to go into more depth but that would be off topic. But Gates talks past all that trying to re-direct the discussion off topic for the sole pupose of attacking on my character.
But several posters saw through Gate’s ugly charade and called him on his dishonesty. So Gates quickly switched to arguing I was misleading people by never discussing the other factors affecting coral. Gates’s lies were again pointed out, showing the essay had 2 sections discussing other factors. Gates just suggested that reality didnt matter and went on trying to create more stawmen.
Gates again switched tactics accusing me of being to over confident when other scientists were not. Gates cites opinions published in 2003 and ignores the growing certainty of many coral experts who are not wedded to climate change gloom and doom. To repeat Dr. Smith observations of reefs like those of the Palmyra Atoll that around uninhabited islands the reef look “almost exactly the same as it did 2,000 years ago. Far from any local human population, Palmyra’s reefs retain 80 to 90 percent of corals, despite past bleaching and ocean acidification.”
The emerging evidence is increasingly detailing the marvelous ability of coral to rapidly adapt, so of course I feel confident about their future. That others disagree is to be expected especially in the highly politicized realm of climate change. But that disagreement does nothing to discredit the Adaptive Bleaching Hypothesis.
Gates then tries to absolve Hoegh-Guldberg by cherry picking quotes and ignoring my links to researchers who stated
Fautin then goes on explain Hoegh-Guldberg’s misstatements.
If Gates was sincerely interested in promoting a better understanding of coral resilience, we would not be “talking past” each other. But as all can see Gates’ mission has been to discredit me simply because my synthesis of coral resilience conflicts with his climate change gloom and doom. He does exactly what he accuses me of doing. He misrepresents the essay, and all that it presents, and tries to divert the discussion to irrelevant details found in cited papers, details that are off topic. Gates is simply one of the more dishonest internet snipers I have yet to meet.
I see nobody has addressed my other question:
Calcification by Reef-Building Sclerobionts by Jennie Mallela PLoS ONE 8(3): e60010. doi:10.1371/journal.pone.0060010
If corals prove unable to adapt, it seem quite possible that brand new complex ecosystems will emerge, which will be a major opportunity for scientific research.
And reduced or reversed calcification (if it happens) will presumably lead to more buffering of anthropogenic CO2, which will help offer time for better technological solutions, without the haste and short-term impacts.
AK,
Let it not be said you don’t have a penchant for finding silver linings. Once again I find myself taking the wrong-by-default pessimistic view that humans presently dependent on current complex ecosystems may not wish them to go away in the name of science. And if I may be so bold, my reading of literature on this topic is that there are yet an overwhelming number of research opportunities.
Not a very convincing argument in that it lacks quantification. As well, more dissolved CO2 in the upper oceans implies a further lowering of pH to which more than just coral species are sensitive. It’s true that over the past 800,000 years atmospheric CO2 concentration experienced swings of 100 ppmv, but at a geologic pace. So this is yet another rate of change argument. We’re effectively in uncharted territory having increased CO2 by 120 ppmv on the order of a century, which is a geological eyeblink.
Jim Steele,
Except Kleypas does not falsify Hoegh-Guldberg (1999). [1] The WPWP is not the whole ocean. That’s a rather important detail to leave out.
As for me being off topic, my understanding is that the topic is corals. Hence, I deem it highly appropriate to discuss other factors affecting coral mortality and/or survivability in addition to relatively rapid temperature change observed *elsewhere* in the world oceans.
Drilling into salient details you’ve glossed over, or pointing out how you’ve misrepresented your own citations is hardly “changing tactics”; I call that critical thinking and appropriate skepticism. Let’s just say that your persistent deluge of ad homimen suggests that you may have little in the way of cogent rebuttal to critique. Please do continue doing it.
———————
[1] Let there be no further “misunderstandings” here. I don’t consider Hoegh-Guldberg the final authority on the fate of corals, or even the main one.
AK says, What if major equatorial coral reefs essentially die? So what? Best evidence is that the general ecosystems will probably move polewards, so it isn’t as though the phenomenon itself will be lost. ”
I personally would be saddened if that happened but I see no evidence to suggest that it will ever happen. A combination of the ocean thermostat and its negative feedbacks, plus due to high humidity and convection the tropics are the least sensitive to rising CO2 accoring to the IPCC, and the fact that the greatest diversity of coral are now in the warmest waters, and that coral have survived warmer waters in the recent past, suggests equatorial coral will be around for a long time.
AK says “And reduced or reversed calcification (if it happens) will presumably lead to more buffering of anthropogenic CO2, which will help offer time for better technological solutions, without the haste and short-term impacts.”
Control of pH in coral reefs is dominated by biology. Photosynthesis raises daytime pH to 8,3 to 8.5 in the summer. In the winter and at night it can drop to 7.5, partly due to reduced photosynthesis
There is also the calcification paradox. The climate concern is lower pH inhibits calcification. But for every mole of calcium carbonate formed, a mole of CO2 and H+ is released and consumption of carbonate ions causes alkalinity to be reduced lowering the waters buffering capacity. Under such self inflicted pH lowering one would expect coral had to have evolved a coping mechanism. Indeed they have and calcification happens in protected vesicles where the coral can regulate the pH.
When pH drops in the winter or at night, exposed skeletons and coral debris can more readily dissolve, but that releases carbonate ions and increases the alkalinity and thus increases the water’s buffering capacity. More and more research suggests that so called acidfication is not a significant concern.
@brandonrgates…
But most of them would probably be willing to take the rather small risk that they go away in the name of abundant, cheap, energy.
This is true of atmospheric CO2, but the exact opposite is true of CaCO3 or MgCO3. Due to the relative solubility of various levels of dissociated carbonic/ates at various pH’s, dissolving these carbonates actually tends to counteract ocean acidification and increasing atmospheric pCO2.
I’ve dug up links for this before, but given that you’re arguing the case, I’m going to leave that as an exercise for you this time. Check it out. (For one thing, there’s quite a few web-sites saying the opposite, and I’m not going to get into a dueling sites situation.) Check out the peer-reviewed science that actually looks at it.
I’d agree we’re “in uncharted territory”, although IMO there are other more significant risks than the effects on coral reefs. And much of the risk, IMO, comes from digging up that fossil carbon and dumping it into the system, even if it ends up in “sinks”. Some of those sinks probably aren’t infinitely elastic.
But AFAIK there’s no reason to consider any of those risks existential, even to our “civilization”, much less our species. This is where the real debate is: do all these possible effects justify urgent efforts to shut down the Industrial Revolution? Or do we have time for technology to solve the problem?
These questions are there in the background of most debates over the science, including this one. It is quite likely that the major coral ecosystems will not disappear with increasing CO2. If they are actually that fragile, there’s no way to distinguish between the effect of CO2, and other more direct issues, such as
• increased sedimentation,
• fertilizer run-off,
• the baleen whales having been driven to (effective) extinction,
• invasive “species” carried in ballast tanks,
• Invasive “species” of barnacle carried by wooden ships,
Etc. This means that even if the dumping of fossil carbon could be quickly ended and/or reversed, there are many reasons to doubt that the corals would return to conditions ante.
And even if the coral reefs don’t recover, the reduced or reversed calcification would actually be a net gain WRT atmospheric CO2.
Is there any clearer evidence that Gates is just trolling and sniping repeatedly tryng to build his bogus case of mischievous omissions . The term shameless evil liar comes to mind.
Here is what I wrote in the essay:
As their evolutionary history would predict, today’s greatest concentrations and greatest diversity of corals are found in the earth’s persistently warmer waters, like the Indo-Pacific Warm Pool. Likewise species inhabiting our warmest waters have undergone the fewest episodes of severe coral bleaching. with a link to the Kleypas paper. that says precisely that.
Now Gates has been called out for his distortion several times now but he just switches lies writing
Perhaps self proclaimed critical thinking does not know the Indo-Pacific Warm Pool and WPWP encompass the same region. What he calls “drilling into salient details” is just hist standard troll drill of creating lies and obfuscation. Surely calling is Gates a shameless evil liar may be construed as an ad hom, but I argue I am just providing salient details about his character that can be verified throughout this comment section. Gates has been caught in so many lies trying to discredit me, there is little recourse but to call a sniping troll a liar when that’s the way he behaves.
AK Part I,
Neither of us need to guess:
In sum, 195 negotiating Parties agree that the best course of action is zero net emissions (not the formal language used) by 2050. Reading between the lines, one *might* say that 195 Parties *assume* the cost of inaction (or business as usual) outweigh the costs of action.
A problem remains that very few countries have actually committed to the 2050 target:
Let’s not confuse what you don’t wish to do with what most of the rest of the world is saying *needs* to be done.
Let’s also not “confuse” my argument here to mean that because most of the rest of the world is implicitly saying that non-mitigation will be more expensive than mitigation that they are right.
AK Part II,
I believe my understanding of the chemistry is the same as yours. Where we differ is that your solution (pun intended) to keeping carbonate concentrations high enough to buffer the oceans to the point that pH remains constant implies that marine species which would normally take the carbonates out of solution for shell or reef building do so at a reduced rate.
I could not have written that any better myself.
Risks not be as extreme as existential to be consequential enough so as to warrant urgent concern.
I think that’s a better question of someone who is actually arging that the Industrial Revolution needs to be urgently shut down. Which isn’t me by a long shot.
I don’t know. Depends on the proposed solution, and how soon it’s enacted. My veiw is that the longer we wait to implement, the worse our odds of costlier and rushed future solutions.
That isn’t what I’m reading in the literature already cited. And here I’m not talking about Hoegh-Guldberg’s estimated worst-case scenario, but of Steele’s other citations which are more supportive of the Adaptive Bleaching Hypothesis. To wit, Maynard et al. (2008):
I think it’s high time for this forum to deal with what that reference and others like it actually say instead of, say, calling me a troll and a liar for pointing it out as Steele has been doing.
Not being a coral expert OR a working scientist of any stripe, I would not presume to declare by fiat what they can and cannot distinguish.
Jim Steele,
Wikipedia defines Internet troll:
The topic is corals. I’ve been talking about corals and environmental issues related to corals, hence I’ve been on-topic.
I am not a coral. Hence talking about me is off-topic.
Brandon Gates writes
Brandon agreed. I would prefer not to talk about you ever. However when you tell lies and fabricate false dichotomies egregiously trying to discredit me, then your bad behavior must be chastised in order to steer the discussion back on topic and maintain an honest debate.
So where are we on t our bet?
I argued that Hoegh-Guldberg’s 95% lost coral by mid-century claim was overly confidence, ill-conceived and really is just fear mongering. Based on evolutionary history, coral’s past history to survive much higher temperatures this past millenium and their amazingadaptive mechanisms, and that climate change is in appropriately being blamed for lost coral, I offered a $500 that coral around uninhabited islands, from the Smith 2016 study, will not lose more than 5% of their coral due to bleaching by the year 2050.
Your history of embracing catastrophic climate change, this should be a bet you would gladly accept. But you seem afraid. Are you fearful that bleaching sensitivity is not due to rising CO2?
Your explanation of why you do not want to bet is your most sincere post to date and well thought out. Indeed I was hoping the “bet” would help focus your thinking. This whole essay is about bleaching and how coral can adapt. As illustrated bleaching has been a very small cause of mortality. And all the media hype, as exemplified by Tripps rants, associate bleaching only with with global warming. I knew if you were to accept or refuse the bet you would need to segregate all the factors. That you converge on a 30% loss due to climate change is far more respectful that Hoegh-Guldberg’ 95% ear mongering. But that still leave’s me wondering why you wont bet that all other things being equal, thus using just uninhabited islands, that climate change will not cause less than 5% loss of coral.
Nonethesless I quibble with your final statement, “that’s one main reason why I’ve been arguing that ABH should not be taken as a virtual guarantee that corals are going to be a-ok.” strays from the facts.
I have never argued that the ABH gurantees the corals will be a-ok from dynamite fishing or cyclones, or pollution that increases sensitivity to temperature variability. When I wrote in the essay “Most extreme bleaching events are associated with El Ninos, but the high mortality rates are not just a function of higher temperatures,” and then I spent a whole subsection on those issues, it was very disingenuous of you to take my comments related solely to temperature and climate out of context, and portray it as if I was ignoring all the other issues.
May I suggest if the obvious was not clear to you, you simply ask for clarification instead of embarking on multiple posts aimed at discrediting me for ignoring those “other factors” that I had clearly mentioned.
May I also suggest that based on your concern about the other factors hurting coral, you could have engaged in a discussion on how do we deal with landscape changes, overfisiing, or any other disruptions. Perhaps then we would have been allies discussing how we humans could make things better. But again you created a false dichotomy, aimed at me personally.
I would have gladly entertained discussion about how to deal with those other factors, even though the point of the essay was to make people aware of corals’ magnificent ability to rapidly adapt to climate change whether it is natural or anthropogenic.
In many of my previous essays I have written about how the erroneous focus on CO2 warming always misdirects our efforts away from the real conservation issues that people can both address and correct ( i.e. walrus, penguins, mangroves, butterflies.) Unwarranted certainty about climate doom often promotes hopelessness that undermines people’s ability to correct the real problems.
In this essay I simply focused on our emerging understanding of how coral can rapidly adapt to climate change via symbiont shuffling and shifting and that usually includes an episode of bleaching. However as alluded to within the essay, there are many other factors affecting corals that have been the primary cause of recent reef declines resulting in a greater sensitivity to El Nino events that would otherwise generate less severe natural bleaching effects. Fear mongering about climate change typically misdirects our attention from those other factors.
A 10 year study, Smith 2016, by coral experts examined and compared coral reefs on islands that are inhabited by people vs those that are not in order to evaluate the effects of human disturbance and the resulting impact on coral resilience. In interviews the principal investigator echoed my confidence in corals’ ability to deal with climate change when free from other disturbances.
“It’s the multiple interacting impacts that give the key to disaster,” says Jennifer Smith, a marine ecologist at Scripps Institution of Oceanography. “If we turned the ocean up by four degrees tomorrow, corals wouldn’t make it. But, if that temperature increase happens over 100 years … I think reefs are pretty good at handling single stress events.”
Smith names the Palmyra Atoll reefs, which she has frequented and studied, as an example of a place that looks almost exactly the same as it did 2,000 years ago. Far from any local human population, Palmyra’s reefs retain 80 to 90 percent of corals, despite past bleaching and ocean acidification.
https://psmag.com/is-climate-change-cooking-our-coral-reefs-58c832a5f631#.6f3s2wuwa
In another interview, “There are still coral reefs on this planet that are incredibly healthy and probably look the way they did 1,000 years ago,” said Jennifer Smith, lead author of the study and a professor at Scripps’ Center for Marine Biodiversity and Conservation. “The scientists were practically in tears when we saw some of these reefs,” she added. “We’ve never experienced anything like it in our lives. It was an almost religious experience.”
http://www.sandiegouniontribune.com/news/2016/mar/20/sd-me-scripps-coral-reef-health/?#article-copy
Such observations from reefs that are free from other human disturbances suggest bleaching events from El Nino events would be relatively minor and would coral quickly recover if not for being stressed already from other disturbances. In contrast to more pristing reefs like Palmyra inhabite island suffer more and make headlines that climate alarmists hijack as evidence of the evil climate change.
I’ll make my bet with Gates even more defined. Of the reefs from uninhabited islands that Smith 2016 observed (as looking just like they did during the warmer waters of the Medieval Warm Period), my bet is those reefs will maintain at least 95% of their coral diversity and cover, compared to Hoegh-Guldberg’s over confident, over simplified and ill informed claim that there will be a loss of 95%.
That wager could too easily result in a tie. That is, you could both be wrong simultaneously (10% loss, for example). So pick one side or the other and specify warming-related bleaching (or similar) to rule out other impacts (major storm, disease, etc.). Besides that, 2050 is a long way off. One or both of you could be dead by then.
Just trying to be helpful. ;-)
opluso,
That was more of a problem with his first formulation:
It’s a sucker bet in *my* favor, not his, because any loss up to and including 95% and I win. Over 95% and he wins. His modified bet includes in part:
This bet only works by him starting with Hoegh-Guldberg’s conclusion of “as much as [up to and including] 95% [of the world’s coral] may be in danger of being lost by mid-century” and mangling it to read “will be a loss of 95%”.
Worse, it’s a sucker bet *against* my favor because now he’s talking about uninhabited islands which presumably don’t carry the same risk factors for overfishing and pollution which were among the significant factors included in Hoegh-Guldberg’s prediction for corals worldwide.
Not only that, but there are conservation efforts underway, some of which may have some hope of success. If I’m reading it correctly [1], Hoegh-Guldberg’s up to 95% was based on a worst-case emissions scenario and little to no other conservation efforts. If emissions were to be reduced, and those conservation efforts were even moderately successful, I’d lose the bet.
———————
[1] Steele pulled the 95% quote from Hoegh-Guldberg (2014), but I believe the estimates themselves come from Hoegh-Guldberg et al. (2007).
First I first saw Hoegh-Guldberg’s 95% in 2014, I never saw it in his 2007 screed. Hoegh-Guldberg does cite his 1999 and 2007 paper but as is often the case his citations dont always back up his assertions.
Second I am more than willing to design the bet so you dont feel at the slightest disadvantage. How much do you feel his 95% loss is due to climate change? 80%? 30%? 1%?
Hoegh-Guldberg did use the lower emission scenarios, But if the world emissions drop lower we can add a caveat that decreases the percentage if emissions fall below projections.
I maintain that observing changes only reefs around uninhabited islands is essential as a constraint because it removes confounding factors from various non-climate factors. The coral bleaching hypothesis can not be applied to other impacts like over fishing, landscapes changes releasing nurtrient etc
Your percentage would also help clarify the issues. How much is climate how much is other abuse factors.
Jim Steele,
I’ve not found anything in the citations I’ve read thus far which estimates that quantity. I previously cited Pandolfi et al. (2003) which says in part:
That suggests the *possibility* that climate change has been responsible for < 50% of observed coral declines up to the time that paper was published, but is silent on the percentage of future declines *could* be attributable to climate change. It's pretty clear to me that Hoegh-Guldberg … discounts … ABH, it's not entirely clear whether he has a problem with the underlying hypothesis itself. Regardless, it *might* be fair to say that he thinks climate change will be the main driver going forward. But I'm only guessing.
That's a long way of saying that I don't have a confident answer to your question.
I also previously cited Descombes et al. (2015) which used the RCP8.5 emissions scenario:
Which nets to 30%. How “reef suitability” maps to “decline in abundance, diversity and habitat structure” isn’t clear to me. Regardless, that conclusion reads as less drastic than Hoegh-Guldberg’s low-emissions scenario estimate of up to 95% loss, and certainly less drastic than the Pandolfi et al. (2003) caution that coral reefs may have on the order of only a few more decades of viability.
I *want* to believe Descombes et al. (2015). I also don’t want to take anyone’s bet — I’d much rather attempt to moot them by actually addressing the risk factors.
Sure, from the perspective of doing the science, it makes more sense to control for confounding factors by looking at environments where those factors are actually absent. From the perspective of placing bets, it doesn’t make sense to me because the mortality projections I’ve been reading are global and attempt to include confounding factors.
That’s my sense as well. At risk of bludgeoning an extirpated equine, that’s one main reason why I’ve been arguing that ABH should not be taken as a virtual guarantee that corals are going to be a-ok.
{This was posted in wrong thread, so reposted)
Your explanation of why you do not want to bet is your most sincere post to date and well thought out. Indeed I was hoping the “bet” would help focus your thinking. This whole essay is about bleaching and how coral can adapt. As illustrated bleaching has been a very small cause of mortality. And all the media hype, as exemplified by Tripps rants, associate bleaching only with with global warming. I knew if you were to accept or refuse the bet you would need to segregate all the factors. That you converge on a 30% loss due to climate change is far more respectful that Hoegh-Guldberg’ 95% ear mongering. But that still leave’s me wondering why you wont bet that all other things being equal, thus using just uninhabited islands, that climate change will not cause less than 5% loss of coral.
Nonethesless I quibble with your final statement, “that’s one main reason why I’ve been arguing that ABH should not be taken as a virtual guarantee that corals are going to be a-ok.” strays from the facts.
I have never argued that the ABH gurantees the corals will be a-ok from dynamite fishing or cyclones, or pollution that increases sensitivity to temperature variability. When I wrote in the essay “Most extreme bleaching events are associated with El Ninos, but the high mortality rates are not just a function of higher temperatures,” and then I spent a whole subsection on those issues, it was very disingenuous of you to take my comments related solely to temperature and climate out of context, and portray it as if I was ignoring all the other issues.
May I suggest if the obvious was not clear to you, you simply ask for clarification instead of embarking on multiple posts aimed at discrediting me for ignoring those “other factors” that I had clearly mentioned.
May I also suggest that based on your concern about the other factors hurting coral, you could have engaged in a discussion on how do we deal with landscape changes, overfisiing, or any other disruptions. Perhaps then we would have been allies discussing how we humans could make things better. But again you created a false dichotomy, aimed at me personally.
I would have gladly entertained discussion about how to deal with those other factors, even though the point of the essay was to make people aware of corals’ magnificent ability to rapidly adapt to climate change whether it is natural or anthropogenic.
Pingback: Den vår de svage kalla höst - Stockholmsinitiativet - Klimatupplysningen
More recent news is here. The Great Barrier Reef just made the headlines.
http://www.huffingtonpost.com/entry/great-barrier-reef-coral-dead_us_574bd64de4b0dacf7ad53513?utm_hp_ref=climate-change
Jim D, Indeed there is ongoing news about bleaching on the reef.
One of Huffpos links reported, “Fortunately, on reefs south of Cairns, our underwater surveys are also revealing that more than 95% of the corals have survived, and we expect these more mildly bleached corals to regain their normal colour over the next few months” says Dr. Mia Hoogenboom, also from JCU.”
South of Cairns also contains the majority of the reefs
http://www.travelstation.com.au/images/map-great-barrier-reef.jpg
Coincidentally as reported in the essay, Osborne’s studies calculated that of all coral mortality bleaching accounts for only about 5%. Coral reefs are dynamics with some reefs declining while others were growing creating a net change of zero.
http://landscapesandcycles.net/image/115377959_scaled_499x240.jpg
Headline saying 93% of the reef has been affected tells us very little, as most of the reef suffered typical mortality of less than 5%. But it does help generate funding. Again bleaching is natural, it is mortality and the lack of recovery that will provide reasons for concern. SO did bleached species become more resilient from one to the next El Nino warm event.
Huffpos headlines, “More Than A Third Of The Coral Is Dead In Parts Of The Great Barrier Reef”
So 2 questions 1) What was the cause for greater local mortality and 2) will the coral recover within a decade as the do from far greater mortality from hurricanes and predator outbreaks?
Warming leads to bleaching, so more warming leads to more bleaching, and the scientists see which direction this is going, which is tough for the Australian government politically, as mentioned in the article.
Jim D
There has also been a rise in cold water bleachings. How do you explain that?
Jim D:
Something like this:
http://myzone.hrvfitltd.netdna-cdn.com/wp-content/uploads/2014/12/General-adaptation-syndrome.jpg
I am going to stick with my theory that the El Nino event will make the coral more resilient to warmer temperatures.
The GBR appears to be warm water because El Nino likely made it worse. Maybe it will recover, but it looks like a downward spiral at current warming rates.
JIm D, It is too simplistic to blame bleaching just on warm waters. There was heavy rainfall and flooding in northern Queensland. Fresh water can be more devastating by disrupting osmosis. How do you separate out the causes of bleaching in the most severely hit regions?
http://www.abc.net.au/news/2016-05-22/record-breaking-may-rain-bom-cairns-weipa-cooktown/7435550
You can look at the historical record. No one recalls anything like this. Warming is the main difference.
come on Jimmy Dee, 2010 had record cold temperatures in the Keys
http://oceanservice.noaa.gov/facts/coral_bleach.html
JIm D, The “historical record” is defined how?
After the 1980s when satellites were used. After the 1950s when scuba was introduced? Again there a historical records of bleaching extending back to the 1800s and bleaching prompted the the Great Barrier Reef expedition of 1928-29 . The two world wars diverted our attention when there was a similar warming peak in the Arctic, Greenland, in ice cores from the Antarctic peninsula instrumental USA data. I would suspect the rapid warming the 1920s to 40s would have been equally stressful but we had no way to observe it in the way satellites allow. There are many researchers who acknowledge this observer bias.
Sure, you can say no way, no how, is it due to the exceptional warming when the scientists have explained why it could be just that with data to support it. I am sure there are ways of aging corals to see how often such things have happened before.
JimD, “Sure, you can say no way, no how, is it due to the exceptional warming when the scientists have explained why it could be just that with data to support it.”
You do realize that if everything was as bad as you think and ALL due to CO2 plus equivalent gases, every imaginable tipping point has been passed. For example there is at least a 10 year lag before emissions and response so by 2026, the globe would be another 0.75 C or so warmer, all equatorial corals would be toast, Greenland would be toast and there would be nothing much more to do that consume all that grain alcohol orally instead of vehicularly.
You and I know that you have no problem overselling in order to inspire policy, but there comes a time when you need to inventory your sales. There really are not that many EMOs that will buy :)
captd, denial of everything just leads to you getting blindsided by these events on an increasingly regular basis. Predict trends ahead instead of steady state. Don’t get blindsided by continuous trends.
“denial of everything just leads to you getting blindsided…”
Jimbo, the biggest denier round here is you, by a considerable margin.
Maybe I can say rejection of everything instead, for the sensitive types.
JimD, don’t know why you think I reject, deny or ignore anything. A doubling of CO2 equivalent gases should cause ~1 C of warming provided all else remains equal. That is down from ~1.5 C per doubling.
Cloud and aerosol impact estimates have also changed over the years and are still a bit problematic. Long term “natural” variability and potential persistence of that variability is just starting to get some renewed interest. I think a recent paper indicated 300 to 1000+ years is the time frame for some variability related to the THC. There should be a few more surprises in store and they should tend to be toward the low side since “believers” have done an outstanding job of selling the absolute worst case.
captd, the problem with lowballing is that you underestimate how fast things change, and get blindsided when they do, so your world becomes gradually more and more unprepared as things progress faster than you continue to expect. This lowballing leads to deficient policies.
JimD, I am not lowballing, I am stating what is actually known. Believers tend to focus on what might be or could be, as if it is known to be.
If I ask you to give the effect of 700 ppm you will lowball and then say nothing to worry about. That is my prediction.
700 from today, 400 ppmv, should result in 0.84 C to 1.5 C of warming, “all else remaining equal.” According to University of Chicago MODTRAN, that should reduce OLR for US standard atmosphere by 2.36 Wm-2. The 0.84 is actual no feedback and 1.5 would be assuming a linear increase, that everything thus far is CO2 equivalent and that there is no increase in negative feedback.
captd, so what does your calculation say for 280 ppm to 400 ppm, and how do you account for the fact that what actually happened was much larger than even your upper ECS estimate of 1.5 C per doubling (requiring a negative imbalance)? In short, your model doesn’t work for the observations unless you invoke a to-be-named-later missing ingredient and figure out what actually cause the positive imbalance of the last few decades. All these questions come from just having lowball numbers.
JimD, “captd, so what does your calculation say for 280 ppm to 400 ppm, and how do you account for the fact that what actually happened was much larger than even your upper ECS estimate of 1.5 C per doubling (requiring a negative imbalance)?”
The simple explanation is that not all of the warming to date is due to CO2. Since “preindustrial” is a tad vague, not that much of a stretch. Since there is a new paper that states the obvious i.e. that the ThermoHaline Current can cause lags of 300 to over 1000 years, persistent warming due to a previous colder spell is now a bit more mainstream. Of course that would mean that someone would have to be wrong *horrors*.
https://lh3.googleusercontent.com/-OvQHzJy8gFU/VLF3XHImAqI/AAAAAAAAMGM/lcW6ogEHxbAukWa0-7MF9Xg7B5Xfx2E_ACCo/s689/oppo%2Bover%2Bmann.png
I doubt that the people wrong will run right out to correct things, but over a few decades things will work themselves out as the old guard loses clout.
You not only have a missing warming component, but also a missing forcing component.
Jim D,
You wrote –
“If I ask you to give the effect of 700 ppm you will lowball and then say nothing to worry about. That is my prediction.”
I don’t know about lowballing (whatever that is), but I ask you to consider that the last time the CO2 concentration was 700 ppm, the surface cooled anyway.
Why is that something to worry about? Has physics changed since the last time the CO2 was 700 ppm?
Maybe you have overlooked the fact that a large white hot ball of rock in a vacuum 150,000,000 km away from the Sun will cool until it is isothermal throughout, beyond the ephemeral influence of insolation – maybe 30 meters or so.
Worry away, if it makes you feel better. Personally, I choose not to. I figure if Warmists were really worried, they’d find a way of doing something about it. So far, they just keep demanding ever more funding.
Paradoxically, after claiming the science was settled, they complained bitterly when the Australian government said “Fine, you don’t need more research money then.”
According to Warmists, the science is settled, unless it affects their hobby. Then, it apparently becomes extremely unsettled. Or maybe it’s just the Warmists becoming unsettled because someone took them at their word!
So, 700 ppm, no problem. If I’m wrong after it gets there, I’ll apologise. Based on history, it will cool again from that point, anyway.
Cheers.
JimD, “You not only have a missing warming component, but also a missing forcing component.”
OMG where! Let’s see the post is on coral bleaching and the overlay on the graphic is for the indo-pacific warm pool which is some of the warmest water in the ocean. The warmest I believe is in the Persian Gulf. According to that and the instrumental for the tropical oceans where a lot of the corals in question reside, there has been uninterrupted warming from the start.
https://lh3.googleusercontent.com/-6Tf2glKTcu0/VG330klOFZI/AAAAAAAALxM/uUtOM1d3aNE8wfSYh5Iq0NfVdsodPW_1ACCo/s800/curry%2Btalk.png
There has been some paleo ocean heat content work done in the same region that indicate the bulk of the oceans have been warming since about 1700 AD or a couple of hundred years before CO2 is supposed to have kicked in. With the exception of the northern latitudes which are extremely noisy, the majority of the globe tends to agree with my estimate.
https://4.bp.blogspot.com/-nfSwpVYkr5s/VzydgFBkLdI/AAAAAAAAOZA/bx-3STUFC_AY0atphqxf5zyFEuvGgOBXQCLcB/s1600/SST%2Btropical%2Band%2Bextratropical%2Bcorrelation.png
JimD, here is another reconstruction for the ENSO 3.4 region using corals that also indicates longer term warming from circa 1750.
https://lh3.googleusercontent.com/-JDDjman7m4I/VHX3peO_mKI/AAAAAAAAL2Y/Y-SzQIIzvcY9KRt7dGrQe_31A38-XtW8gCCo/s679/nino%2B3.4%2Bemile-geay%2B2012.png
Additional CO2 will no doubt have some impact but additional evaporation and convection will offset a large portion of that impact, in the warmer water regions which make up a little over 50% of the oceans. Since “other” stresses are mentioned for just about every warm water coral region being damaged, I imagine focusing on things that can be addressed in a reasonable time frame is a bigger bang for the buck than something that might take centuries and likely won’t have any significant impact. Odd that the coral paleo seems to indicate more than just CO2 isn’t it?
Jim Steele (May30, 5:01PM) poses the
So 2 questions 1) What was the cause for greater local mortality …”
I gave the answer for this above (May 29, 2016 at 2:37 am). And as I wrote there, more can be seen at my site.
pj I will check out your site later today
Test
Note that the hypothesis of this post is currently being investigated:
Climate change disables coral bleaching protection on the Great Barrier Reef
Under the BAU (high carbon) scenario (RCP 8.5) the GBR is doomed and under likely impossible to accomplish dramatic reduction scenario (RCP 2.6) the GBR is probably doomed as well, so focusing on other sources of stress, COT, pollution, etc.is recommended.
I imagine that if you are an advocate of extremely high CO2 forcing impact, putting your head between your legs a kissing the GBR goodbye is the way to go. However, if you are a tad more optimistic of the future, trying to save the GBR might be worthwhile, starting with things that can be changed on a realistic timescale.
Pat there have been several papers since 1993 investigating the adaptive bleaching hypothesis. The Ainsworth 2016 paper you link does acknowledge adaptation but do not acknowledge symbiont shuffling and shifting,
So the authors acknowledge corals’ ability to rapidly adapt. So why do they speculate that model projections of global warming will “disappear” their adaptive mechanisms?
This paper does not take in to account the effects of symbiont shuffling and its effects on tolerance limits and assumes a rigid phyiological response. The paper only assumes that a “pre-stress” event (slightly warmer than average maximum temperatures) prepares coral for future stress events (greater than 2C warmer than average maximum temperatures) that result in bleaching. They then predict that global warming will metamorphose pre-stress events into stress events. But that assumes no symbiont shifting
However if coral can acquire more heat tolerant symbionts as has been increasingly observed, corals’ heat tolerance limits rise, and their physiological adaptive mechanisms remain in place but for a new set of temperatures.
MAP: Mass bleaching has killed a third of the northern Great Barrier Reef
http://edge.alluremedia.com.au/uploads/businessinsider/2016/05/coral-bleaching-map.jpg
JCH,
Global warming has now become Cooktown warming by the look of the graphic.
According to the link, this is the third time in 18 years that this has happened. I presume that twice before, the reef recovered. Why not this time?
Maybe something else is responsible. There seems to be less bleaching at places further towards the Equator, where SSTs are hotter.
What’s your explanation? It all seems a bit inconsistent, to say the least,
Cheers.
2/3rds of coral survives unprecedented high temperatures.
Lol. 2/3rds of family survives deadly car crash.
Seems rather odd that global warming unleashed its swath of hig percentage death only at Cooktown. Seems like other events may have been involved like flooding
http://www.abc.net.au/news/2016-05-22/record-breaking-may-rain-bom-cairns-weipa-cooktown/7435550
“The coastal area that I study north of Broome has huge tides, and we thought the corals there are tough “super corals” because they can normally cope with big swings in temperature,” says Dr. Verena Schoepf from the University of Western Australia. “So, we’re shocked to see up to 80% of them now turning snow-white. Even the tougher species are badly affected”.
https://www.coralcoe.org.au/wp-content/uploads/2016/04/GBR-Results-LowRes-700×923.jpg
JCH,
Do not confuse temporary bleaching with mortality. And reports of 93% of the reefs bleaching does not convey what percentage of a reef bleached or how temporary.
The most recent reports are south of Cairns mortality has been less than 5% of the coral.
I didn’t say anything.
Call the scientists who study coral and warn them to not get confused.
JCH,
So it looks like global warming only greatly affects corals north of Mackay. Odd warming. Seems localised to me, and to the scientists, apparently.
Is there a particular reason that there would be greatly increased CO2 levels north of Mackay? How were other natural cyclical or quasi cyclical coral bleaching episodes ruled out?
I suppose you’ll claim lack of knowledge. It seems that Jim Steele is prepared to do a bit more research than your hasty cut and paste of prima facie illogical and contradictory graphics.
There are peer reviewed papers indicating that symbiotic relationships can, and often do, exhibit dynamical or chaotic population behaviour. Throw in the odd predator prey relationship – say Crown of Thorns starfish infestation – and chaos is pretty much guaranteed.
I don’t mean to rain on your parade, but not every facet of Nature can be ascribed to burning filthy, dirty coal. Panic as much as you want, but remember that more than 99% of all species that ever existed on Earth became extinct before the influence of humans.
Why waste a good worry?
Cheers.
Pssst. It grows back. Otherwise we wouldn’t have had coral to do any dying in ’83 and ’98 after the severe COTS hits of the ’70s (which lasted eight years on the northern reef).
Those tour boats haven’t been making their money showing where coral used to be. They’re showing lots of live pretty coral today.
The greatly increased population of activists and professionally concerned people has certainly been bad for tourism in Qld, especially now that El Nino has done its bleaching thing again. But what’s to be done? You can’t just inject sodium bisulphate solution into climate warriors like you do with the COTS.
mosomoso
I predict your mosomoso will lose its leaves in the next 40 years and or flower. This time-unlike the other times through history-It will be due to AGW. Do something about it NOW.
Don’t look to me for help as I will be occupied trying to prevent leaf fall on our oaks this autumn which will be due to man.
tonyb
It’s amazing. Cat 5 Cyclones like Mahina in 1899 (world record for surge height, if you’re into records) and Innisfail 1918 (end of Innisfail Mark 1) made a mess all on their own. Now a Cyclone like Yasi can look just like Mahina and make a similar mess…but it’s man made!
Not a lot of people know that. Just the highly educated ones who read the Guardian and HuffPo.
You can’t be too paranoid these days.
https://climatenuremberg.files.wordpress.com/2015/09/dear-god-no-b.gif
H/t Climate Nuremberg.
Looks like Glenn Beck’s scruffy older brother.
‘ A new syndrome Munchhausens by proxy describes
the use of swamp varves and tree rings to pretend
the earth has a fever, typically in hopes of obtaining
sympathy, attention or other secondary gain.’ )
Jim Steele,
[Posted out of sequence for scroll.]
No, because
a) that would be good news and
b) it’s inconsistent with my reading of published literature.
Here’s what Smith et al. (2016) has to say in the abstract (I can’t find the full document):
That suggests other direct human impacts (like fishing, agricultural runoff etc.) are the dominant mode of coral mortality. From the PS Magazine article you cited:
She’s giving 10% as her low-end bound for coral mortality in the Palmyra Atoll reefs, so a 5% bet *for those reefs* might make sense.
Elsewhere in literature, bleaching events from temperature excursions are described as a *compounding* factor contributing to coral mortality, loss of diversity, etc. Very next paragraph of the same article, the previously mentioned Baum and Cobb:
So the $500 question in my mind is how long can we expect the recovery to take? Recall Baum and Cobb saying in the previously cited article that one reef they looked at twenty years on from the 1998 El Nino was only just beginning to show recovery.
My statement is consistent with what two of your citations — Baker (2005) and Maynard et al. (2008) — say about ABH.
In your essay you cite McClanahan et al. (2015). Searching for a full copy of the paper, I came across a full copy of McClanahan et al. (2011) which also invokes a multivariate analysis approach that does not mainly rely on temperature thresholds. I thought the Introduction and concluding paragraph were interesting:
I have no further comment to add to those excerpts, they’re consistent with arguments I’ve already made numerous times above.
With all due respect, when you cut and past lengthy abstracts , it is never clear what point you are trying to make. I suggest more concise replies and a more focused reply would make for a much better discussion.
Jim Steele,
I find it not always easy to excerpt literature in a concise way that preserves context. But I can, and arguably should, summarize my point(s) when including a long copypasta as I have done above. I’ll begin with my final citation, McClanahan et al. (2011), and work backward. This section is most relevant to the point I wanted to make:
Makes two interesting points:
1) Temperature anomalies in and of themselves are not good global predictors across *all* reef systems.
2) Studying heterogenous temperature responses may give insights into how we might enhance coral resilience (and/or adaptation) to climate change.
I’ve touched on both points previously, this reference combines them in a way that … feels … right to me. My additional comment is that we would do well to remember that bad OR good news in one reef, or a system of regional reefs is not necessarily representative of *all* reefs globally. OTOH, it’s not wholly unreasonable to suppose that commonalities do exist, e.g., that global temperature excursions might be expected to have (nearly) globally adverse effects with something like ABH being one “built in” mechanism giving hope for (nearly) global recovery after a widespread temperature excursion-induced bleaching event such as we have witnessed in the 1998 AND 2015 “Super” El Nino events.
*IF* it turns out that large El Ninos “ratchet up” absolute temperature thresholds for most coral populations with little to no further intervention on our part, that would indeed be good news.
The Smith et al. (2016) abstract suggests that human factors other than rising ocean temperatures *may* be dominant. The PS Magazine about the article included counterpoints from her colleagues Baum and Cobb, however Smith herself makes a good one:
The cautionary note I’m seeing is that multiple impacts compound … as in they’re not simply additive. That dovetails into this:
Whereas I see it as a question of what *combination* of actions can we take to minimize global coral mortality, or stated in the positive, to conserve their resources to our own benefit. Everything I’m reading suggests that both global and regional/local actions will tend to maximize their chances. OTOH, the global solution — reducing CO2 emissions — will be a long-time coming, and is slow-acting … temperature rise would not stop on a dime even if net emissions were brought to near-zero tomorrow. Thus we *probably* should put an emphasis on developing strategies to enhance coral adaptation/survivability, part of which includes further studying ABH for ways that we might enhance it in a directed and locally-targeted fashion.
I do not mean to imply that reducing emissions cannot or should not be done in parallel.
***
I don’t think that necessarily follows. Lamy (2016) Fig. 1 identifies specific events associated with observed declines at Tiahura reef, namely COTS and tropical cyclones. Going back to McClanahan & friends’ work on multivariate models, it *seems* plausible to me that *compounding* factors can be isolated.
And just to be clear, I would personally be hesitant to ever suggest that a dead reef got that way 100% due to CO2-driven climate change.
Brandon Gates says
That suggests other direct human impacts (like fishing, agricultural runoff etc.) are the dominant mode of coral mortality. From the PS
Yes of course, as discussed in the essay, bleaching is the least cause of mortality. Cyclone damage and human impacts that favor coral predators or stress coral account for 95% of coral mortality.
Brandon Gates says,
“So the $500 question in my mind is how long can we expect the recovery to take?
A most excellent question that I would expect from a sincere truth seeker. The graph of reef destruction and recovery from Lamy 2016 in the essay suggests that 15 years is a minimal amount of time required to determine if a reef is in the midst of a cyclical mortality/recovery period or trending downwards
http://landscapesandcycles.net/image/115377991_scaled_616x310.jpg
So what is the natural succession withing coral communities? Terrestrial plant communities have adapted to natural fire regimes. Several pines do not release seeds from their cones until there is a fire that created plenty of sunlight. Many species have seeds that will not germinate until scarred by fire. Other plants are pioneer species that quickly invade open areas cleared by fire. Their seeds also require high solar irradiance, but their parents shade, prevents them from persisting. Shade tolerant species begin to flourish and after 20 to 50 years begin to dominate the forest until the next fire starts the cycle again.
There is no reason to believe coral reefs do not undergo similar cycles that require 50 or more years to complete. Like fire adapted plants, there are fast growing corals that are quick to colonize after destruction from a cyclone or predator event. Fleshy algae are also quick to dominate after widespread coral mortality from cyclones, depredation or natural bleaching, and can out-compete coral for space. Normally fish populations that graze on fleshy algae begin to increase denuding the reefs of seaweeds so that coral regain their foothold.IF overfished that process is reduced. However there are locations where fleshy algae dominate immediately afters a disruption, but die off during the winter, allowing coral to rapidly regains dominance
So the $500 dollar question depends of several confounding factors, and that’s why I insist that the bet be constrained to reefs around uninhabited islands that are not overfished. In pristine places bleaching will still happen but recovery should be rapid. On average I would suggest there recovery would take no more that 10 to 15 years in more pristine reefs. If recovery is not progressing then we can call the reef dead, and suggest a high likelihood of climate change.
I hope all the aerial surveys were followed up by divers checking for predators such as Crown of Thorns starfish.
“The Crown-of-Thorns Starfish (COTS) is a natural predator of corals in the Indo-Pacific region, including the Great Barrier Reef (GBR). COTS have been identified, together with cyclones, as a major cause of 50% decline in coral cover on the GBR observed during the last 27 years (De’ath et al. 2012). Since the 1960’s, COTS populations have erupted at approximately 15- year intervals with three major outbreaks recorded and a fourth now in progress on the northern GBR. When COTS occur in plague proportions they can reduce the living coral cover on a reef to a few percent. Full recovery from such severe disturbances requires at least two decades free of all major disturbances (e.g. cyclones, coral bleaching, more COTS, e.g. Halford et al. 2004; Emslie et al. 2008). The causes and consequences of COTS outbreaks have received much research attention since the mid-1980 and a recent review by Pratchett et al. (in press) summarises the current state of knowledge.”
All part of Nature’s rich tapestry of life.
Cheers.
Thanks for a common sense artical. Much of the scientific speculation about destructive or abnormal or increased coral bleaching is about getting funding for research. The coffers are wide open from envirnmental groups, governments, and others, for promotion of anthroprogenic climate warming, but not for honest research.
Brandon Gates quotes the Descombes paper and their model that suggests global warming will reduce coral by 46% in their current locations.
http://doc.rero.ch/record/257087/files/pel_fcr.pdf
However their study has several flaws and their model does not take into account symbiont shifting. They only reported, ”
That laboratory study is 25 years out of date with the new understanding of symbiont diversity and heat tolerant clades and species. HOwever they do report coral with higher tolerances and the ability to acclimatize
They also use the Eocence as an analog for future climate assuming strong CO2 sensitivity but oddly to model the Eocene they use 2250 ppm CO2. Furthermore fossil samples from the Tethys Ocean are used to suggest higher temperatures in the past had reduced coral diversity. But the Tethys was slowly closing as Africa and India pushed northward, currents were changing, volcanic activity was high, and shallow seas were developing anoxic conditions. Low diversity can not be simplistically blamed on higher temperatures. But they generated a curve based on average annual temperatures observed and modeled to suggest coral diversity reaches an optimum between 28 and 30C, Thus any climate models projecting higher temperatures also predict lower coral diversity and abundance. The logic behind their 46% loss is based on that curve suggesting “suitability for coral reefs strongly decreases in SST conditions warmer than approximately 32° C”
It is also not clear if the ocean thermostat hypothesis was included in their models of future tropical ocean temperatures, a hypothesis that suggests warm pool waters are much less sensitive to heating which has resulted in less coral bleaching. See Kleypas http://onlinelibrary.wiley.com/doi/10.1029/2007GL032257/abstract
Jim Steele,
They give a nod to the possibility:
Palumbi et al., 2014 is relevant.
An open question for me is whether modelling ABH is feasible at this point in time. Although a somewhat dated reference, Buddemeier et al. (2005) has this to say:
I assume progress has been made over the past decade. My question is whether enough has been made to develop a *global* model which takes ABH into account.
Reads to me that they assumed whatever sensitivity and warming is implied by the members of the CMIP5 ensemble for RCP4.5 and RCP8.5.
Wikipedia tells me that CO2 concentrations during the Eocene aren’t well constrained (700-900 ppmv to upwards of 2,000 ppmv depending on the proxies used), so 2,250 ppmv doesn’t seem wholly out of bounds. It’s also worth noting that Descombes & Co. didn’t actually do the Eocene climate model, but obtained the model results from Huber and Caballero (2011), who actually also proposed a 4,480 ppmv CO2 scenario for the Eocene, about which they say:
A careful reading of the entire paper (or at least the preceding paragraph or two) to put that comment into proper context is suggested. I infer that Descombes et al. used the EOCENE-2250 model because the coral fossil record for the warmer early Eocene is much sparser.
More salient to this discussion, my reading is that Descombes et al. did more than just use the middle Eocene as an analog for future climate …
… more that they took a beyond the worst-case scenario by 2100 to better characterize the response curve to temperature. Note: I would not have written “full response curve” because I’m a bit of a stickler about not implying that “we” know everything there is to know. ” … thereby more fully modelling the response curve … ” would sit better with me.
My reading is that they suggest that conditions in the Tethys Sea went past an upper threshold unsuitable for corals. Like you, they list a number of other factors which may also have contributed to the dearth of corals in the Tethys during the Eocene:
I would also point out that their conclusions were NOT based solely on conditions in the Tethys Sea. If I’m reading the paper correctly, they used fossil records representing 204 reef systems, about which they say: During that period, coral fossil records were well represented in the western Tethys, eastern Pacific and Atlantic oceans, but in very low frequency in eastern Tethys characterized by high mean annual SST up to 36.7 °C (Huber & Caballero, 2011)
We’ve come full-circle to Kleypas et al. (2008), which was focused on the WPWP. A key point to consider comes directly from the abstract …
Paragraph [16] puts a finer point on my argument here:
I assume that Descombes et al. evaluated the forward-looking AOGCMs they used for fidelity to observed SST evolution; the paper suggests their selection was based on a number of factors including ocean chemistry. So the extent that they considered an ocean thermostat mechanism in warm pools would be implicit in the extent that the AOGCMs they used correctly project SSTs for a given emission scenario. I’m of the mind that the only way to know that for certain is to wait and see what happens.
Which is, of course, not exactly my favorite option.
http://notrickszone.com/2014/07/04/caribbean-coral-reef-die-off-not-caused-by-climate-change-after-all-expert-report-writes/#sthash.nNHBW091.dpbs
The lead author says words to the effect of, Climate Change is “an excuse for doing nothing”
Even doing something like putting up a windmill in Minnesota isn’t going to help the coral reefs.
“The decline is not uniform and correlates only weakly with local extreme heating events, instead being mainly attributed to the severity of local stressors, in particular tourism, overfishing and pollution.”
I don’t know how they could say such a thing? Weakly correlated?
matthewrmarler,
[posted out of sequence for scroll]
Good grief, Matthew. Steele used the phrase “virtual guarantee” in comments several times.
By definition, there can be no evidence of future events. What we do have is evidence that rapid temperature excursions (both cooling and warming) can cause widespread bleaching and thence widespread coral mortality. From the paleo record, as shown in e.g. Descombes et al. (2015), we see that there *may* be an upper temperature threshold beyond which corals cannot adapt. Hence it is *plausible* that sufficiently high upper ocean and surface temperatures *could* result in degradation/loss of low-latitude reef systems, especially *if* the *rate* of temperature change is high relative to more geological time scales.
*Maybe* ABH will tend to minimize the damage we’ve already witnessed from temperature excursions AND other factors like lowering ocean pH, overfishing and pollution from land runoff. My position is that literally betting the fish farm on it isn’t a Good Idea.
There were no guarantees in the essay. My “virtual guarantee” was a comment in context of a discussion of thriving coral and their resilience to warmer temperatures in the recent past, a past inundated with more frequent El Ninos.
Brandon used that one comment several times and juxtaposed it to other researchers’ uncertainty in order to launch several attacks on my credibility and redirect the discussion.
To add clarity to my stance. I stand by my virtual guarantee that based on historical evidence (2.1C warmer in the holocene optimum) and adaptive bleaching, that if all else was equal, the coral are not threatened by the current rates of warming whether or not that warming is natural or attributed to CO2.
But indeed all things are not equal. Damage from landscape issues, sediments, pollution, dynamite fishing, cyanide fishing, over fishing, damage from tourists and the aquarium trade, boat anchors, building of channels that redirect the currents, etc all add considerable uncertainty as to how well the coral will respond during normal bleaching events, and how rapidly they can recover. Researchers pushing CO2 warming as the main issue argue all those events are secondary, but it is equally correct to argue current warming is secondary to warming. As the essay stated, bleaching is the smallest contributor of coral mortality.
The solution is to manage reefs locally where we can control directly things like setting up no take fishing zones. The Thai governnent was wise to shut down reefs to tourists, because heavy tourist use interferes with the normal recovery period to a bleaching event.
brandonrgates: . Hence it is *plausible* that sufficiently high upper ocean and surface temperatures *could* result in degradation/loss of low-latitude reef systems, especially *if* the *rate* of temperature change is high relative to more geological time scales.
With respect to the climate change debates, I am a great fan of “maybes”, and if that is a summary of your opinion, I agree with you about the plausibility of possibilities. .
Many possibilities are plausible. Random variation and natural selection are among the best documented phenomena in science. It is more than just “plausible” that every species of coral has some offspring in every generation that will have greater reproductive success at higher temperatures than they do now. It is especially “more than just plausible” given that extant coral species have survived higher temperatures than contemporary temperatures in the past few thousand years.
“overfishing” and “pollution” are not the reasons given for the calls for us to imminently reduce fossil fuel consumption. That is based on the threat that extra CO2 will produce warming. So warming is a good topic to focus on.
Steele used the phrase “virtual guarantee” once. I can “virtually guarantee” that whatever new medicine is introduced this year that has a terrific success rate in treating TB, in 20 years there will be resistant TB in every geographic region in which the new medicine is used. There are already glyphosate resistant weeds in America’s farmlands. In both cases, more than 95% of the target populations are killed in the first few generations, much higher than coral mortality from warmer weather..
matthewrmarler,
Not for the first time, I bring up the Big Five mass extinction events. Here’s what Veron (2008) has to say about those events w.r.t. corals:
We could quibble about whether Veron pinning the greatest *potential* risk on carbon cycle changes and ocean pH really is the most *plausible*, but we don’t have to for the message to come through: corals’ adaptive mechanisms have been overwhelmed five times in geologic record and they have been wiped out, not to return for *at least* four million years according to this reference.
Possibility with known precedent five times over is the sort of plausibility that I can’t easily dismiss on the basis that “many possibilities are plausible”. YMMV.
I count five, which is consistent with my description “several”:
Note that the first usage of “virtually guarantee” was in response to my challenge on the broader and less qualified prediction that corals “possess mechanisms that will allow them to adapt to any warming in the near future”. This is all in stark contrast with what ABH’s proponents say in references Steele himself provided in support of his own position:
Baker (2003):
Maynard et al. (2008):
Shifting and shuffling away from what the words on the page actually say does not help your case, Matthew.
brandonrgates: Possibility with known precedent five times over is the sort of plausibility that I can’t easily dismiss on the basis that “many possibilities are plausible”. YMMV.
Which of those 5 extinction events is most like the gradual increase of global mean temp to a value that was met several times in the last 5 millenia? Are you now focusing on the likely change of mean ocean pH of about -0.3?
The prospect of ocean acidification is potentially the most serious of all predicted outcomes of anthropogenic carbon dioxide increase.
Or will corals benefit from the increase in CO2 as have the coccolithophores and other marine organisms that have been studied?
Ah, I see that you are correct on the count of “virtual” guarantees, which you shortened to “guarantees”. I stand corrected.
brandonrgates: Indeed, there is growing evidence that such shuffling can increase thermal tolerance, at least in the short term (Berkelmans and van Oppen 2006). This is a promising area of research, but heralding symbiont shuffling as a panacea for reefs threatened by climate change is premature.
I don’t make claims about “panacea”. I wrote that random variation and natural selection are among the best documented phenomena in science. So adaptation of a large population of small, rapidly reproducing organisms such as coral to changes of about +2C in temp and -0.3pH is much more plausible than their extinction. That quote refers only to a few possibilities among the many possible random variations. Given our experience with viruses, staphylococci, spirochetes, mosquitoes, and weeds, I would bet that there are a lot more possibilities than that.
brandonrgates,
I see that it was I who shortened “virtually guarantee” to “guarantee” . I apologize.
Brandon Gates is lapsing into more disingenuous posts. It was he that made the “virtual guarantee” an issue, not by discussing any evidence that would support or refute such a comment, but by juxtaposing it to comments by others who were less certain. Gates repeated the same tactic over and over and then when it evoked another response then he tries to portray that comment as if it was something I was repeatedly pushing. But that is the nature of Gates dishonest personal attacks that misdirects the issues.
As I said if all else remained equal, then coral should thrive in warming trends that repeat the natural warming of Holocene Optimum with temperatures 2.1C warmer than today.
A sincere and scientific disagreement would offer evidence and theory why it might not be true that coral could survive similar warm temperatures. But Gates never does that. His posts were less about the science and more about trying to shoot the messenger. Gates simply tries to re-direct the issue by arguing if another scientist is more uncertain then that somehow their uncertainty invalidates my argument. And he trollishily did that over and over again.
So if Gates is trying to be more sincere now, then perhaps he can provide a more meaningful argument why coral will not once again survive in oceans that 2.1 C warmer than today!
And I suggest we need a more substantial argument than Tripp or Hoegh-Guldberg’s that the rate of warming is happening at a much faster rate than ever before. That facts are simply not their for tropical waters. Again the Nino 3.4 region shows NO trend. And it is the redistribution of warm water events by El Nno that has triggered these widespread bleachings
https://i2.wp.com/data.giss.nasa.gov/gistemp/2011/Fig8.gif
Jim Steele: Brandon Gates is lapsing into more disingenuous posts.
I think that the scientific evidence that has been cited strongly supports your opening essay. However, it was my mistake to shorten the phrase [virtual guarantee] and then to claim that no one was arguing thus. I am embarrassed by it.
Brandon Gates argues
Gates seems to be unaware that the word coral refers to very different evolutionary lineages. The relevant lineage for today are the Scleractinians that were only around for the last 2 mass extinctions and had just gained prominance during the Jrassic before being decimated by the asteroid impact. To be clear, I have never argued that symbiont shifting and shuffling has ever prepared coral for an asteroid impact that cause the last great mass extinction, and why Gates thinks it may be relevant quite mysterious.
Veron also presents temperature data that also supports the ocean thermostat hypothesis. Despite speculation of high CO2 during the Mesozoic Veron reports minor differences in tropical waters
Furthermore while bleaching is restricted to coral with a symbiotic partnership (aka zooxanthellate corals) Veron reports that during the most recent mass extinction, the K/T,
Again Gates’ post is irrelevant. The extant lineage of coral that have survived today are different from the extinct corals with adaptive mechanisms that were overwhelmed five times in geologic record. But Gates is not a coral experts so such details were likely not obvious.
matthewrmarler,
Here’s a global temperature reconstruction for the past 11.3 millenia after Marcott et al. (2013):
http://www.realclimate.org/images/Marcott_s3.jpg
Here’s the abstract:
The study doesn’t support the contention that *rate* of change over the past century is unprecedented in the past 11.3 millennia, but they don’t rule it out either. Where’s your evidence that *rate* of warming since the mid-20th century is “gradual”?
As I have been from almost the beginning of my participation on this thread, I’m attempting to focus on as many identified risks to coral reef viability as I can find, falling ocean pH being one of them I’ve mentioned previously.
My main argument at present is that the Big Five extinction events are strong evidence indicating that ABH has indeed failed to keep corals from getting wiped out, and thus should NOT be relied upon as a virtual guarantee of their future survivability. As a mechanism which *may* be a useful feature which gives hope to our own ongoing preservation efforts, yes certainly. But not a get out of jail free card when it comes to CO2 emissions.
I’m going to go with what refereed literature I’ve cited says, Matthew, including the references Steele cited in support of ABH. I’m not a corals expert or a working scientist.
Neither will I insist that Veron (2008) *must* be correct — I’m not into single paper syndrome. I also can’t do all your homework for you. I know that annoys the hell out of folks when I say it, but here I do insist — these issues are too complex for me to do all the heavy lifting in terms of research and critical thinking. I will not be able to “prove” to you that my position is correct.
Yes, Steele’s first four instances were “virtually guarantees”, so searching on “virtual guarantee” wouldn’t find them. Stuff happens, not a big deal, and no need for apologies — though I do accept yours with thanks.
Yes, I realize that.
Which I don’t dispute.
That doesn’t *necessarily* follow. You might be correct, but the thresholds you’ve assigned have no obvious direct relationship to the relative rapidity with which corals reproduce, which you have not quantified. I’m not inclined to accept your argument because at least one reference specific to corals and prior mass extinction events suggests that corals’ adaptability is bounded, and well short of boiling the oceans. Look, my sense is that even the coral pros have difficulty and disagreements amongst them about the best way to approach understanding (and predicting outcomes for) what is a fiendishly complex dynamic system. If I have a main overall point to make, that one would be a good candidate.
Regarding the sense that ABH implies adaptation by way of evolution of corals themselves, Buddemeier et al. (2005) have some interesting things to say:
Perhaps a bit of a quibble, but there it is. I also think the first sentence of that paragraph is worth pondering.
brgates, Just curious, how much stock do you put in the accuracy of Marcott et al. ? Personally, I was a tad curious and found a few higher frequency reconstructions of temperature in the same areas and often by the same authors, that would extend the lower frequency reconstructions he used into the 20th century instead of ending in the 15th or so. I also just used the tropical reconstructions, a few of them based on corals.
https://lh3.googleusercontent.com/-3k7UcdrOfRU/VLgkkI95IPI/AAAAAAAAMKU/vsje2z28wysjLbeEXM4w0tq-rGcERjiFwCCo/s676/tropical%2Bholocene.png
brgates, in case you up to a critical thinking challenge, the Marcott et al. 2013 supplemental data can be found here.
http://science.sciencemag.org/content/suppl/2013/03/07/339.6124.1198.DC1
Many of their reconstructions have “caps” or reconstruction made using the upper portion of a core which is typically done with a different analysis method since top isn’t as compacted as the majority of the core. For whatever reason, Marcott et al. didn’t include these “caps”, so they make pretty fair out of sample data. You can find many of caps at NOAA paleo.
If you like Marcott et al. 2013 “as is” and think it is stellar paleo-oceanography, here is your award :)
https://wattsupwiththat.files.wordpress.com/2014/09/comic_rollercoaster_610.jpg
Jim Steele,
The Veron quote supports polar amplification, not warm pool thermostasis. The key is that the temperature change differentials are expressed in terms of zonal means.
Revisiting Kleypas et al. (2008):
In sum, *if* the hypothesized WPWP thermostat breaks, corals in that region *could* be in for a rude surprise.
You’ll note that I’m not trading on my personal expertise, but the expertise of those in the literature I cited to make that point.
FWIW, I’m fully aware that extant “corals” are thought to have come from different lineages than extinct “corals”, it was discussed upthread and I was responsive to that discussion. Problem with your argument here is that you’re prevailing on the *possibility* that extant corals are different from extinct species. Why corals should have evolved (or re-evolved) to better handle temperature excursions after the final of the Big Five extinction events isn’t at all clear to me.
Here, let’s review what the very first reference you cited in this essay, Baker et al. (2008) has to say:
Seriously, most of the points I’ve been arguing on this thread are right there in that one abstract. Here’s what you had to say about that paper: As one expert wrote, the “flexibility in coral–algal symbiosis is likely to be a principal factor underlying the evolutionary success of these organisms”.
I don’t need to be a coral expert to understand what coral experts themselves actually say about their outlook, nor our pressing need to intervene on their behalf for our own common good.
brandonrgates: Marcott et al. (2013):
We critiqued that when it first came out. Its problems are serious.
Well you know, Dallas, I like Marcott et al. (2013) better than Marler’s non-cited reference to Holocene global mean absolute temperatures.
Here’s what the highlights for Hertzberg et al. (2016) have to say:
Full paper available here.
lol, ok. Tree rings are the highest frequency proxy I know of for the past several centuries. Can’t really beat annual resolution.
There are probably reasons why studies like MBH98 and Marcott et al. (2013) use multiple proxies. Just sayin’.
If there aren’t any further squirrels to chase, let me thank you for your input.
matthewrmarler,
Everybody and their dog critiques every proxy reconstruction when it comes out, including the referees. But this is getting tedious. Here again is your question to me:
That question implicitly asserts that the temperature rise in the past century is “gradual”, and present absolute global mean surface temperatures have been met several times over the past 5,000 years. So. You’re not happy with Marcott et al. (2013), which is fairly consistent with your claim, albeit silent on rate of change. Fine. Cough up a reconstruction which does support your claims. Thanks.
brgates, “If there aren’t any further squirrels to chase, let me thank you for your input.”
You are a piece of work, all those tex86 and g. ruber reconstruction are used by Marcott et al :) They use a Tierney et al TEX 86 and totally screwed the dating along with not using the cap. so I reckon if the proxies Marcott et used are crap it must be crap :)
The g. ruber mg/ca cold bias btw depends on region because of currents and while not perfect beats the crap out of uk’37. There are some coral reconstructions like Emile-Geay 2012 that happen to agree pretty well with those worthless g. ruber mg/ca proxies.
https://lh3.googleusercontent.com/-JDDjman7m4I/VHX3peO_mKI/AAAAAAAAL2Y/Y-SzQIIzvcY9KRt7dGrQe_31A38-XtW8gCCo/s679/nino%2B3.4%2Bemile-geay%2B2012.png
captdallas,
In conjunction with other proxies. That’s why I was talking about multi-proxy studies in my previous post to you. Dig? Here’s a hint: the plots you posted based only on those two proxies alone *might* not give the most robust results, eh?
Speaking of being a piece of work, I notice you ran away from the higher resolution is more better argument when I invoked tree rings.
Says who? Please don’t bother with a citation that hasn’t been published in a refereed journal.
And you know what? Go bang on Marler for a credible source supporting his assertions about GMST over the past 5,000 years so that the discussion of how that may or may not effect corals can move forward.
brgates, “In conjunction with other proxies. That’s why I was talking about multi-proxy studies in my previous post to you. Dig? Here’s a hint: the plots you posted based only on those two proxies alone *might* not give the most robust results, eh?”
Combining similar proxy reconstructions, same methods, frequency and “surface” (ocean-ocean, land-land) should have a more predictable bias than a blend different proxies and regions that have different and unpredictable biases. All reconstructions based on biological organism that desire to survive will have some bias to an “optimum” range.
https://lh3.googleusercontent.com/–sQaqVGQEVo/VKloPB1CnQI/AAAAAAAAMEs/edyZ3rq6AYUdiUkqk7TIN07kJZ2NEcTQwCCo/s757/Tropics%2B4000%2Byears%2Bindividual.png
As for the dating issue, that doesn’t have a peer reviewed report but it is easy to check and has been noted by quite a few folks, Nick Stokes is just one. There are a fairly large number of reconstructions that have been revised thanks to online critiques and post-publication reviews.
captdallas,
The best possible scenario would data going back to the beginning of time taken from modern (or better) quality instrumentation.
Well ok you found a bias; I’d entertain Stokes as a reference on this one. Gotta citation?
Which sounds like good science to me. Now back to the original question; what’s a suitable reconstruction that substantiates Marler’s claims about GMST over the past 5,000 years? I submit that Marcott et al. (2013) is consistent with his claims, but both you and he don’t like it because “its problems are serious” in his words.
Or … were there more squirrels yet to chase? I don’t want to miss anything truly salient to the corals issues we’ve been discussing here.
brgates, “Which sounds like good science to me. Now back to the original question; what’s a suitable reconstruction that substantiates Marler’s claims about GMST over the past 5,000 years? I submit that Marcott et al. (2013) is consistent with his claims, but both you and he don’t like it because “its problems are serious” in his words.”
Probably Rosenthal et al. 2013 discussed here a while back, https://judithcurry.com/2013/11/01/pacific-ocean-heat-content-for-the-past-10000-years/
Remember though that has a resolution of about 50 years tops so you would need to be careful how you compare that with instrumental. Something like this.
http://4.bp.blogspot.com/-UxfO3016ARs/UpvcTaKSfmI/AAAAAAAAKqw/LS89ybhG5PI/s1600/2000+years+of+climate.png
Here is that link to Nick Stokes discussion of Marcott. He never bothered with the caps and other available data so I did that myself.
http://moyhu.blogspot.com/2013/03/my-limited-emulation-of-marcott-et-al.html
brgates, According to the new v4 version of ERSST, there should have been a major bleaching event around 1890 :)
http://climexp.knmi.nl/data/iersstv4_115-120E_-3–6N_n.png
This tropical cooling could be problematic
captdallas,
Fanks. Press release declares:
I dunno, is it an exception to Betteridge’s law of headlines when the subhead (sorta, kinda) answers the question? On the plus side, I see that Mann didn’t overwhelmingly endorse it. Anywho, a pre-print of the paper is available here. Imma go right for the pretty picture, Figure 2:
https://2.bp.blogspot.com/-Far6IodKtYI/V1HdjNaIPyI/AAAAAAAAA3w/EsEcK2foglEgiu4owSgTIYGOiYYS2SENwCLcB/s1600/Rosenthal%2B2013%2BFigure%2B2.png
Compare Marcott (2013):
https://i2.wp.com/www.realclimate.org/images/Marcott_s3.jpg
Pretty similar if you ask me.
Thanks again. He writes in the head post:
The complaints are mainly about some recent spikes. My main criticism of the paper so far is that they do plot data in recent times with few proxies to back it. It shows a “hockey stick” which naturally causes excitement. I think they shouldn’t do this – the effect is fragile, and unnecessary. Over the last century we have good thermometer records and don’t need proxy reinforcement. Indeed the spikes shown are not in accord with the thermometer record, and I doubt if anyone thinks they are real.
I agree with all he writes there except the final sentence: it was not immediately apparent to me that the spikes coinciding with the modern instrumental period were not in accord with the modern records, and I was rather rudely surprised to learn about that some time after the paper had been published.
Other than that, he doesn’t find issue with the balance of the reconstruction that I can see, and it seems to compare well with Rosenthal. What am I missing? Oh …
If this whole discussion weren’t about trying to substantiate Marler’s claim that recent warming is “gradual”, I’d be more inclined to dig into that right now.
captdallas,
A 5×3 degree grid near the equator hardly constitutes “the tropics”:
https://3.bp.blogspot.com/-njTGxkKs_A4/V1HmlQEx4NI/AAAAAAAAA4M/218t0WmvrSoEhPz-5VTsukk9LVPRjyCgwCLcB/s1600/iersstv4_0-360E_30–30N_n_a.png
Here’s Hadisst’s take:
https://3.bp.blogspot.com/-i23q_GP40go/V1HmlVn_IAI/AAAAAAAAA4Q/viZdw5z5BSsVLRU3EK_LEOzqiGmt4Mz5QCLcB/s1600/ihadsst3_0-360E_30–30N_n.png
Whut? Dialing down the “Demand at least: x % valid points in this region” option from the default 30 to 5:
https://4.bp.blogspot.com/-S6zU9tOnVWU/V1HmlTYkrCI/AAAAAAAAA4U/U_y07LjTXgITCxmOMxkg2v6D9eUC08d1wCLcB/s1600/ihadsst3_0-360E_30–30N_n_5p.png
Especially back that far, does the problem of relatively small number of observations not occur to you?
brgates, How about a bit bigger area?
http://climexp.knmi.nl/data/iersstv4_110-146E_0–16N_n.png
I believe the top part of the GBR is in that area. BTW, those pictures you like happen to be from Marcott 2013 and include the spurious uptick, one of the issues with Rosenthal et al. The ERSSTv4 has been created since the publications of Marcott et al. and Rosenthal et al. which creates a bit of a problem for the either the paleo community or the community that uses 1880-1899 as a “pre-industrial” reference :)
The part of Rosenthal et al. that is interesting is the 300 plus years of gradual warming they show once you get past the PR hype.
brgates, “Especially back that far, does the problem of relatively small number of observations not occur to you?”
Of course it does, compared to paleo though instrumental is a couple of orders a magnitude larger. That is a major issue with Marcott et al. because adding the caps and other reconstructions available indicates their confidence interval is a tad optimistic. Using ocean proxies with the huge ocean inertia helps, but 10,000 years with a +/- 0.15 C uncertainty is almost unbelievable :)
btw, here is some of that pesky text that relates to your brilliant comparison of Marcott to Marcott :)
“To the extent that our reconstruction reflects high-latitude climate conditions in both hemispheres, it differs considerably from the recent
surface compilations (i.e. Marcott et al. 2013), which suggest ~2°C MWPto- LIA cooling in the 30°N to 90°N zone, whereas the 30°S to 90°S zone warmed by ~0.6°C during the same interval (24). In contrast, our composite IWT records of water masses linked to NH and SH water masses imply similar patterns of MWPto- LIA cooling at the source regions The inferred similarity in temperature anomalies at both hemispheres is consistent with recent evidence from Antarctica (30), thereby supporting the idea that the HTM, MWP, and LIA were global events.”
captdallas,
I posted my own already.
So it is.
That would tend to essplain the eerie similarity and my more than usual confusion about these things — I’d asked for a global MST reconstruction and you feed me a paper that does SST reconstructions for the Makassar Strait and Flores Sea. No excuse for me to not have read more carefully, but I take it you understand my dilemma.
At least we agree that the uptick between 1890-99 is likely spurious.
I don’t suppose you have an example of this problem relevant to corals?
I confess, I haven’t gotten that far yet as I’ve been rather more focused on things I know to be directly relevant to the corals discussion. Did you know that NOAA hosts a corals listserver?
Ove Hoegh-Guldberg lays out why he thinks the ABH is wrong.
Daphne G. Fautin defends her hypothesis.
Bob Buddemeyer, Fautin’s husband and co-ABH developer basically says Hoegh-Guldberg is strawmanning the hypothesis.
Interesting reading the arguments from experts outside the constraints of refereed literature. Lotsa other fascinating stuff in that thread.
“I don’t suppose you have an example of this problem relevant to corals?”
Only Emile-Geay et al. which used ersstv3 for part of their paper but I focused more on g. ruber recons. However, calibrating to a moving SST target is going to have impacts when the move is significant.
http://journals.ametsoc.org/doi/abs/10.1175/JCLI-D-11-00511.1
Apparently the Australian government has produced a (secret) report that claims it will cost $16 billion to meet water quality targets over the next decade to help save the Great Barrier Reef.
http://www.abc.net.au/news/2016-06-02/great-barrier-reef-pollution-fight-could-cost-billions/7469740
It gets worse… It also states that the total estimated cost of delivering “all sediment abatement investments” could in fact be in the order of $33 billion, while nitrogen abatement could top $12 billion.
Who should we believe? A blog essay that claims this bleaching event (magnified by anthropogenic CO2 global warming) is just a minor setback for the coral and that, quote: “Symbiont shuffling and shifting is an evolutionary masterpiece that circumvents plodding evolutionary mechanisms of most organisms with long generation times and enables immediate adaptation.” or the government of the sovereign nation of Australia. Well Jim could be right. If the Australian government did spend the billions of dollars to restore the water quality then it may indeed be possible for the symbiont to adapt to a warmer climate. I’m going to go out on a limb and bet that the money won’t be spent and the reefs will be pretty much gone in a few decades, mostly due to pollution but greatly assisted by global warning.
Yep, kind of a waste of time to fix the easier stuff when the whole system is doomed.
jacksmith, You conflate very different issues. Adaptive bleaching and symbiont shifting is a marvelous mechanism that has allowed coral to survive climate change. That’s what this essay is about. It doesn’t mean it will help coral survive when a boatload of sediment smothers the coral or if it instigates a crown of thorns outbreak.
Juxtaposing those different issues just creates a misleading and disingenuous argument.
I acknowledged the thesis that symbiont shifting can adapt to warmer oceans, I just doubt humans are willing to make the effort to give them the chance to evolve fast enough to restore the ecosystem to it’s original state. Do you think Australia should spend the 16-33 billion to stop the pollution? I don’t. Will spending that money improve the standard of living of the average Australian? Not much in my opinion. There will always be a place somewhere where coral will survive, right?
brandonrgates,
You quoted –
“Surface temperature reconstructions of the past 1500 years suggest that recent warming is unprecedented in that time.”
The non Warmist translation might be “We say our guesses are better than your guesses, so there!”
Species die out. New ones emerge. At least, according to history. As to the future, who knows?
The Earth’s surface has apparently cooled over the last four and a half billion years, the supposed greenhouse effect notwithstanding. As the continents move around, rising and falling as they go, habitats change. The marine fossils above 6000 m in the Himalayas demonstrate that ammonites were unable to adapt to changed conditions. So what?
Peer reviewed guessing competitions notwithstanding, trying to scare people into removing proven plant food (CO2) from the atmosphere doesn’t seem like the smartest thing to do for the continued existence of our species.
I understand your passion. As far as the Earth heating up due to CO2 in the atmosphere, you’re just as wrong as Kelvin was about the age of the Earth. Newton’s belief in alchemy was widely shared, but now searching for such things as the Philosopher’s Stone seems irrational. He was wrong.
And so it is with Warmism (or whatever sciencey name you wish to ascribe to it) – passionately espoused by believers. Irrational and somewhat bizarre, in common with most cult beliefs.
Richard Feynman referred to the phenomenon of Cargo Cult Science. Climatology seems to fit his criteria very well.
My assumption is that the universe is unfolding as it should, warts and all. Assigning a care factor above zero to increasing CO2 levels, seems to me to be a complete waste of time. Obviously, your mileage may vary!
Cheers.
Mike,
One nasty habit of experts is that they tend to believe they know more about their own domain than non-experts. Ridiculous proposition, I know.
Wake me up when conventional lay wisdom routinely improves brain surgery outcomes.
Jeeze, what is it with the squirrel-chasing today?
Food crops did just fine at 280 ppmv, though modern fertilizers and herbicides/pesticides surely helped a great deal. The LIA was perhaps a mite coolish, word is that Goldilocks lives somewhere about 350 ppmv. But I don’t pay those arguments much mind because I think they’re a moot point — I believe we’ll be lucky to stay under 550 ppmv by 2100. So have no fear, I don’t think we’re even remotely close to being able to starve plants by taking away their CO2.
My mileage does vary. I’m particularly amused that your appeal to directed outcomes follows right on the heels of invoking Feynman’s musings on Cargo Cult Science. That kind of flip-flop would make my brain explode … I’m frankly amazed that yours hasn’t.
brandonrgates,
I’m not sure to whom you are referring as experts. Climatologists are presumably experts in calculating averages, as climate is no more and no less than the average of weather. Any reasonably competent 12 year old possesses the same level of expertise. Definitely not brain surgery, or even rocket science.
As to trying to use the worn out Warmist tactics of attempting to deny, divert and confuse by including climatology in the same continuum as brain surgery, you lose, as usual.
Brain surgery is useful, on occasion. Climatology, on the other hand, has produced nothing of benefit to the human race.
I’m not sure what your squirrel reference is about. Another deny, divert, confuse attempt?
You may not like real scientists (doing replicable real experiments based on real plants and real CO2) pointing out that the world is greening, and that plants tend to grow more prolifically when fed better, but that’s real science for you.
Your unsubstantiated musings about an optimum CO2 level are just silly. It’s about as silly as claiming that deserts are hotter because they have more greenhouse gases over them! That would be really silly, wouldn’t it?
Do you really believe that Hansen, Mann, Schmidt, Trenberth and the rest actually know what they’re talking about? I guess you do, otherwise you wouldn’t have written some of the things you have.
As Trenberth once said “It’s a travesty!” Have fun!
Cheers.
Wrong!
AK,
Per IPCC AR5 –
“Climate
Climate in a narrow sense is usually defined as the average weather, or more rigorously, as the statistical description in terms of the mean and variability of relevant quantities over a period of time ranging from months to thousands or millions of years. The classical period for averaging these variables is 30 years, as defined by the World Meteorological Organization. The relevant quantities are most often surface variables such as temperature, precipitation, and wind. Climate in a wider sense is the state, including a statistical description, of the climate system.”
From World Meteorological Organisation –
“Climate is generally defined as the average state of the atmosphere for a given time scale (hour, day, month, season, year, decade and so forth) and generally for a specified geographical region.”
Climate normal per WMO –
“A Normal is defined as the arithmetic average of a climate element (e.g. temperature) over a 30-year period.”
Maybe you can provide a contrary definition?
Or do you think that to a Warmist, climate is anything the Warmist declares it to be at the time? That would certainly be convenient!
Cheers.
https://judithcurry.com/2016/05/20/week-in-review-science-edition-42/#comment-785472
Not that I insist on my definition. But nothing anybody can do can change the “average of weather”. Anyone claiming that something is “changing the climate” is using a different definition, functionally. No matter what they claim.
The “average of weather” is an artifact of our limited knowledge, and at best is something we can look at to
measureestimate the state of “the climate”.And yes, it is a semantic quibble. But semantics are important. When people use words with “slippery” definitions, they tend to talk past one another.
Mike,
Someone published in primary literature probably didn’t get there completely by mistake. There are notable exceptions … I personally like picking on Pons and Fleischmann as the epitome of previously undetected cranks. And there are a slew of lower-tier journals with, shall we say, more permissive publication standards. And it’s not like Nature has never had to do a retraction.
I think that more or less covers my bases.
When the average 12 year old can build a complex climate model and write the documentation for it, again, please wake me up.
Doesn’t matter what I write, Mike. If it contains the words “climate” and “CO2” within the same paragraph, you ain’t gonna believe it. I’d have better luck herding cats.
The former is a metaphor for the latter. As in interjecting the old “CO2 is plant food” chestnut into a conversation about corals.
I think that kind of research is just fine. I’m also fond of pointing out that CO2 helps noxious weeds grow too … that conclusion comes from the same papers.
Term I used was “moot”, but silly works.
That’s your argument, not mine, so of course the answer is yes. Fun fact: the Sahara and Amazon have roughly the same mean annual temperature, but the diurnal range is much larger in the Sahara. My own fellow American, Bill O’Reilly, might triumphantly say, “You can’t explain that!” I don’t know what the Ozzie equivalent is.
BTW, you’re chasing squirrels again. Or perhaps more accurately at this point, Gish Galloping.
They make more sense to me than you do. Not that I set the bar *that* low as a general rule, mind, but I’m just sayin’.
The best time to look for the greenhouse effect is night. The deserts get colder at night, Mike. What do you make of that? Headscratcher, eh? Quite a puzzle. Just another one of the world of mysteries to people who don’t understand physics.
AK,
The simplest definition I recall reading is that climate is statistics of weather over some arbitrarily long period of time. As that could be the mean, median, min and max, standard deviation (the “classic” descriptive statistics) of any given parameter, I don’t find it incompatible with your definition — which I quite like.
As for the shape of the attractor, well yes, I understand that to be an issue especially with all the known and supposed non-linearities existing in such a complex system … to which you allude below.
Perhaps I misunderstand this argument, but I’m having trouble visualizing how geological-time external forcings a la Milankovitch would become inseparably smeared in with internal modes of variability like AMO or ENSO.
Geological-scale modes of internal variability beggar belief save for, e.g., continental movements which have an effect on ocean circulation.
Come on. Over the last century the CO2 signal is dominant and clear as a bell. CO2 went from 280-400 ppmv; the system response has been unequivocal and more or less in line with Arrhenius’ predictions near the turn of the 19th Century. Since you don’t think so, I think it’s incumbent on you to propose a different mechanism tied to a discrete set of physical principles and model it. Beating the CMIP5 ensemble at its own game would be particularly convincing.
brandonrgates,
1. You still decline to nominate your supposed climatological experts. Your
choice.
2. The CAM 5.0 model, like other climate models, produces results which differ from reality, according to peer reviewed evaluations. Predictive ability no better than a 12 year old with a ruler and historical data.
3. Weeds? A weed is just a plant you don’t like!
4. Assume a CO2 level of 350 ppm. What golden age will ensue? No floods, droughts? Will the climate will remain fixed and immutable for ever? No new temperature or precipitation records? You really can’t say, can you? What’s the scientifically based (not according to CAM 5.0, for sure!) reason for your chosen CO2 concentration?
5. Fun fact : average temperatures are completely pointless for useful climate or weather comparisons. In fact, several members of a British SAS patrol died in Iraq, because someone assumed an apparently comfortable average daily temperature was a useful indicator of clothing and equipment requirements. The patrol members who died of hypothermia might have benefited from the advice of a local 12 year old child, peer reviewed or not.
6. Michael “I talk to trees” Mann actually knows what he’s talking about? Are you sure? Treemometers? Maybe you should check with Gavin Schmidt.
So it looks as though non existent warming due to the presence of CO2 in the atmosphere is not going to kill much coral anytime soon. I’ll let you have the last word. Enjoy.
Cheers.
Jim D,
Of course deserts get colder at night. Basic physics – little water vapour in the atmosphere, and little cloud. Greater radiative loss to space. That’s also why the hottest places in the world are found in arid tropical deserts. Less impediment to insolation reaching the surface.
No puzzle to me, just basic physics. I suppose post normal Warmist physics claims that deserts are hotter due to some bizarre greenhouse effect. Oh well.
Cheers to you too.
Mike,
I have a known resistance to answering tedious questions. Here’s an answer you’re sure to like: lead authorship for an IPCC WGI chapter is probably a good indicator of eminence within that particular domain.
All models are always wrong, Mike.
Unless you’re 11, I look forward to you besting it. Gridded, all output variables, from 1861-present. Chop chop.
You’re right, I don’t much like eating weeds. Eating is important. One might call me addicted to it.
Given the lags in the system, my expectation would be rather similar today albeit a tad cooler.
Don’t be silly.
No seriously, stop being silly.
You’re a one-man strawman machine.
I have a better chance of characterizing the familiar than I do the unprecedented.
I told you it was a moot point, but 350 ppmv CO2 coincides with the year 1988. We know for a fact that year was compatible with modern technological civilization and five billion in human population.
Fun fact, you glossed right over the bit about diurnal range and rambled off chasing yet another squirrel. Color me shocked … well wait a tick … you actually answered Jim’s point about nighttime temps: Of course deserts get colder at night. Basic physics – little water vapour in the atmosphere, and little cloud. Greater radiative loss to space.
Next you will fail to explain why CO2 is an exception to the same rule.
I’d wager he’s forgotten more about paleoclimate than you’ll ever be interested in learning.
I thought it was Tom Wigley who set Mann straight on the unsuitability of using tree rings as a temperature proxy.
Your leaps of logic are indeed dizzying.
I always do enjoy toying with you. Character flaw on my part, I’m sure.
Below is the abstract from Silverstein Change in algal symbiont communities after bleaching, not prior heat exposure, increases heat tolerance of reef corals
As I have presented in this essay the focus is on how symbiont communities can be shuffled and shifted an increase coral resilience. No one in the scientific community would accuse them of misleading anyone about the other stressors that coral face. This contrasts with attempts by several posters here to divert the issue of increased heat tolerance.
The points made here are precisely what this essay has argued.
Let’s review the abstract
Furthermore acquisition of D1a symbionts is an example of coral acquiring symbionts that were “pre-adapted” to higher stress level.
Jim Steele,
At least one poster here thinks a main problem with the essay and your subsequent comments is that they over-egg the pudding. And I’m sorry, but attempting to consider the full suite of risk factors facing corals isn’t a diversion from the issue of coral survivability — the very papers you cite in support of ABH without exception make at least some attempt to put their findings into that context. They certainly don’t suggest as you did in comments that there are any guarantees, virtual or otherwise, of survivability in the face of expected future warming.
Abstract is essentially the same as what we’ve already been shown. Paper itself again suggests that there are no guarantees, and includes this interesting note on why corals literally should not expect any free lunches as a matter of course:
To be fair, further down we read:
Given the choice between dead and suboptimal, I’d choose suboptimal for sure. But I wouldn’t like it that those were my only two choices.
Something else Silverstein et al. (2014) wrote caught my eye:
The full reference is Hoegh-Guldberg O, Mumby PJ, Hooten AJ et al. (2007) Coral reefs under rapid climate change and ocean acidification. Science, 318, 1737–1742. The much-derided H-G (1999) lurks in the references as well.
Anyway, I thought the 2007 paper was an … interesting … citation in this context, and it confused me a bit until I reevaluated the “underappreciated” modifier. Here’s what H-G et al. (2007) say about ABH:
Clearly not ABH’s biggest fan is Dr. Hoegh-Guldberg, but I wouldn’t call that passage worth being nailed to a post for heresy against it. To their credit, Silverstein et al. didn’t — I presume in no small part due to the fact that their respective conclusions are strikingly similar in most other respects:
Again, pretty much what I’ve been arguing about and pointing to in your own citations all along. That I question ABH’s *potential* efficacy w.r.t. warming in the context of *all* other potential global risk factors is something that its own proponents do, thus one wonders to whom your leading complaint …
… should actually be directed.
The paper supports most of what I have been arguing.
The title of the essay was “Coral Bleaching Debate Is bleaching the legacy of a marvelous adaptation mechanism or a prelude to extirpation?”
And Indeed the paper echoes my stance that adaptive bleaching allows adjustment to climate change saying,
And it also shows Gates’ lack of understanding about this rapid adaptation mechansim. Brandon Gates remark about the pepper moth was a false analogy and just an excuse for unfounded snarkiness. Inappropriately throwing in phrases like “punctuated equilibrium” was again more evidence of his lack of understanding of how corals’ adaptive mechanism works
In Silverstein’s experiment adatpation was observed in just 3 months Clearly they show that bleaching is “the legacy of a marvelous adaptation mechanism”
PS: I would be remiss to not note that when H-G et al. (2007) say this about ABH …
… they are misstating the mechanism of ABH. It isn’t the generation times and genetic diversity which are the operative mechanism, but the shifting and shuffling of symbionts within a given coral’s own lifetime. So I warrant they do deserve to be dinged for at least that much about ABH in this passage.
In reply to my statement regards the Silverstein paper “No one in the scientific community would accuse them of misleading anyone about the other stressors that coral face.”
Brandon Gates wrote
My comment was aimed at Brandon to point out why many of hiss comments were inappropriate.Like Silverstein I pointed out in my essay that there were other factors affecting coral mortality but the focus of ths essay was on the Adaptive Bleaching Hypothesis and that it provides evidence for coral resilience to climate change. In order to do that the essay had to take a more narrow focuse muck like Silversein does. TFOr Brandon to argue there are many other factors affecting coral survival was something I clearly stated and do not disagree. I was simply pointing out that Brandon’s tactic tried to denigrate the essay by implying a narrow focus was an attempt to hide other factors. Such attacks are typical of snipers not scientific discussion.
Jim Steele,
Again, here’s Silverstein focused solely on climate change:
Compare your two previous statements in comments above:
I don’t think it at all inappropriate to point out that what you write is not compatible with the references you cite to support them. Your citations propose a mechanism which may/might/could allow *some* corals in *some* reef systems adjust to higher sustained SSTs. You issue confident predictions and virtual guarantees.
Sixth time’s a charm isn’t a very compelling argument. I think it imprudent to discount past mass extinction events due to rapid environmental change on the basis that *some* extant species *might* be better able to cope with expected future change than previously thought — especially when the species in question are already showing rapid decline due to a number of combined and likely compounding factors.
Per WUWT, a debate rages in Australia over additional funds to improve water quality/pollution sources near the GBR. Closing the pollution barn door after the climate change horses have escaped?
Original source is subscription only.
https://wattsupwiththat.com/2016/06/03/expert-scientists-exaggerated-coral-bleaching-story/
a) The existence of other threats to corals does not in any way suggest that bleaching is not a threat. The phenomena are separate and independent.
b) The argument that an adjusted ecosystem will survive is undoubtedly correct but also completely irrelevant. 20% of fish species live in the reefs as they currently are. While many of these species may be able to adapt to the new conditions and new species may emerge this process will take decades or centuries during which ocean productivity will be adversely impacted. A large proportion of the world’s population is dependent on marine agriculture for their food security. Large-scale effects that undermine global food security are not negligible.
If bleaching is a generalized stress response, it’s probably not much of a threat. Perhaps an indicator of a threat…
Well, if enough refuges survive, most of the important “species” probably will too. Meanwhile, if the reefs don’t come back in their current form, whole new ecosystems will proabably emerge, with large number of currently rare “species”.
Hm… This could be a problem. What proportion of that “marine agriculture for […] food security” is actually dependent on reef productivity? As opposed to, say, open ocean plankton?
And BTW, how much of it is really “agriculture” as opposed to mechanized hunting/gathering? And would you consider mariculture to be threatened by issues with natural reefs?
Anthony says
“a) The existence of other threats to corals does not in any way suggest that bleaching is not a threat. The phenomena are separate and independent.”
No one has ever argued that other threats mean beaching is not a threat.That would be silly. Regards “separate and independent” phenomena the reply is depends. Tropical storms are the biggest natural threat totall independent of how coral will adapt to warmer water. But cyclones do cause bleaching coral recovery mechanisms that evolved to deal with storm mortality provide the same resilience to bleaching from other causes
Yes, corals have recovery mechanisms, these mechanisms apply against all forms of external trauma. This is irrelevant to whether or not the currently observed bleaching episodes are a cause for concern or not. To suggest that AIDS is not a problem because septicemic plague is worse would be equally fatuous.
Anthony all you are doing is posting false equivalncies
You are quite correct, the harvesting of seafood is economically more similar to open cut mining than any form of sustainable activity.
Mariculture has demonstrably been seriously impacted by climate change. There have, for example, been particularly adverse effects on abalone and oyster production in Tasmania. These species are particularly dependent on depth and rising sea levels have further required relocation and adaptation of infrastructure.
More intense storms and higher storm surges have impacted coastal activities of all stripes but particularly mariculture, aquaculture and low-lying agriculture.
Brandon Gates linked to the Veron paper to argue coral did not survive the big 5 extinctions events. However most of the coral that went extinct were not related to today’s coral families and therefore the point was irrelevant. One of the more interesting points however, Veron also presented temperature data that also supports the ocean thermostat hypothesis.
So while the global average may have undergone large temperature swings, the tropical waters did not. The negative feedbacks that inhibit tropical warming suggests coral may not be subjected amplified warming from CO2. Even the IPCC writes the tropical waters are the least sensitive to the greenhouse effect.This why Kleypas pointed that coral living in warmpools have undergone the least amount of bleaching
However Brandon Gates tried to dismiss that evidence but only revealed once again he is just slinging contrariness and again revealed his lack of understanding, writing
Polar amplification is a hypothesis tha the poles will lose ice due to rising CO2, and the lost ice reduces albedo resulting in warmer temperatures. BUt te Veron quote was from the K/T period when there was no ice on Antarctica or in the Arctic.
No Ice, No polar Amplification
The difference between tropical and polar temperatures during that time was due to changes in how warm tropical waters are transported poleward and has nothing to do with polar amplification. And Ocean Oscillations that pump more warm water poleward is likely the best explanation for the decline of Arctic sea ice in the 1930s and in the past decade.
Jim Steele,
I erroneously included a partial response to this post above; I continue that response here in its proper sequence.
See Barron and Washington 1985 Figure 2, which is consistent with polar amplification. That paper does not give any support for a thermostat mechanism that I have found. It’s the next sentence in Veron that cites evidence of a thermostat mechanism:
I think Veron overstates how widely accepted the ocean thermostat hypothesis is, which has its roots in Ramanathan and Collins (1991), Newell (1979), Hartmann and Michelsen (1993), later extended by Li et al. (2000) and Clement et al. (2005). Fu et al. (1992), Wallace (1992), Pierrehumbert (1995), Sud et al. (2008) and Williams et al. (2009) all challenge the notion that there’s an effective global upper SST limit. Here’s a sampling from Williams et al. (2009):
I also think Veron (2008) overstates the case for a thermostat as made in Kleypas et al. (2008):
Both papers suggest that a global SST thermostat may be an untenable hypothesis.
Payne et al. (2015) only partially agree with you:
Thus …
… isn’t supported by all literature. Also note that CO2 is not the only external forcing where surface albedo feedback leading to polar amplification would apply; solar output and/or orbital forcing (a la Milankovitch theory) would be expected to have a similar effect. In the case of orbital forcing, the surface albedo change could be seen as both a driver and a feedback.
Yet changes in poleward heat transport would be expected to affect the amount of polar amplification in a regime of global temperature change.
… but silent on — and rather at odds with — what would explain the unabated increase in global OHC over the past several decades.
Brandon says, “I erroneously included a partial response to this post above; I continue that response here in its proper sequence.”
That’s total BS. You imply didn’t know what you were talking about and engaging in your normal dishonest snipes. Call it a partial answer to cover your butt, but have witnessed your tactics far too often to eve believe that.
This is too funny. Brandon cuts and pastes whatever abstract he finds hoping to find something thta will stick. First he snipes at my citation to Kleypas citing the fewest bleachings in the warmest waters, because I didnt delve into the ocean thermostat’s ability to minimize warming. As Veron stated tropical waters warmed very little relative to the global average polar regions. Such small changes in the warmest waters is support of some sort of thermostat process which is backed by solid physics. The Clausius Clapeyron relation shows at a certain temperature, increasing heat flux increases evaporation rather than heating the water. A combination of increased evaporative cooling and reduced insolation act as negative feedback.
Noooowww Brand wants to tell us Veron overstated his case? LOL And First Gates overstates the connection to coral extinction confusing extinction causes and totally unrealted reef builders. Gates also fails to complete Verons evaluation
Veron stated
Hmmm CO2 levels were 5 to 10 times that of today and temperatures of the largest oceans would have been limited by the Thermal Cap of 31C.
That seems like more evidence to back my virtual gurantee.
Indeed polar amplification in its simplest terms is the much greater warming in the polar region than the global average. Amplification has only happened in the Arctic, and a small region of the Antarctic. With the context of CO2 driven climate change, it is the belief that CO2 will melt the ice and snow and reduce albedo.
Regards coral survivability it is changes in tropical waters not global averages that is important. And As the IPCC states CO2 has only a small effect on tropical temperatures, the CO2 theory points to polar amplification as a function of less ice more heat.
In contrast and like myself, many researchers point to the Arctic amplified warming as a function of inatural oscillationss and ntruding tropical waters, lost thick insulating sea ice due to Arctic winds, resulting in more heat ventilating from the ocean.
Although Gates cites Chylek as a researcher who provides evidence against the polar amplification as merely a function of melting ice, Gates fails to accurately detail Chyelk’s paper that explains high Arctic temperatures in terms of natural oscillations. CHylek wrote
Jim Steele,
Because Kleypas explicitly states that Ocean thermostat mechanisms do not appear to work effectively everywhere in the tropical oceans.
You are aware that coral species are found outside the WPWP, are you not?
Not necessarily, see again Payne et al. (2015).
For crying out loud, no:
Clausius–Clapeyron equation
(Also called Clapeyron equation, Clapeyron–Clausius equation.) The differential equation relating pressure of a substance to temperature in a system in which two phases of the substance are in equilibrium.
There is no time component in the differential equations listed, therefore there is no way to solve for a flux quantity.
Read again the opening clause of first sentence of the block you quoted: Just what that would have meant for reefs is uncertain …
Yes, about the wide acceptance of the ocean thermostat hypothesis.
Try Chylek et al. (2010) on for size:
[5] In the following we show that the multidecadal scale variability of detrended Arctic and Antarctic temperature time series were highly anticorrelated during the 20th century. Consequently we suggest that the Atlantic Ocean may provide the physical link leading to the coherent multidecadal scale climate changes of opposite phases in the two polar regions. We further suggest the ocean variability contributed at least as much to the recent Arctic warming as the increase of atmospheric concentration of greenhouse gases (GHGs).
[…]
4. Summary
[17] A bipolar seesaw pattern of the paleo temperature has been observed earlier in the Greenland and Antarctic ice core data. For the first time we identify a bipolar seesaw pattern in the 20th century Arctic and Antarctic instrumental temperature records. The detrended multidecadal scale variability of the Arctic and Antarctic temperature time series are highly anticorrelated. When the Arctic warms Antarctica cools and vice versa. This multidecadal variability combines with the general warming trend (presumably dominated by anthropogenic GHGs) to produce the observed Arctic and Antarctic temperature patterns. The intense Arctic warming since the 1970s (Figure 1a) arises from an additive combination of the general global warming trend with the warming phase of the multidecadal climate oscillation, while in Antarctica the cooling phase of the multidecadal oscillation opposes the general warming trend leading to essentially no significant Antarctic temperature change since the 1970s (Figure 1b)
brgates, “For crying out loud, no:
Clausius–Clapeyron equation
(Also called Clapeyron equation, Clapeyron–Clausius equation.) The differential equation relating pressure of a substance to temperature in a system in which two phases of the substance are in equilibrium.
There is no time component in the differential equations listed, therefore there is no way to solve for a flux quantity.”
C-C works well up to 100% RH then not so much. As the air approaches saturation, that would be 100% RH, you start needing something a bit more advanced than C-C, like kick butt cloud and convection models. Since model assume a constant RH but C-C indicates an increase in RH (it is an ideal model that assumes water is always available), there is a bit of an issue with a lot of “super” adjectives, super-saturation, super cooled liquid. This is great because other guys like Antoine get equations named after them. Sharing the scientific wealth and fame.
brgates, btw, whether or not you need a revised version or replacement for C-C depends on how well it is estimating what you are trying to estimate. OMG! You cannot figure it out because there is no time dependent variable in the equations! doesn’t really cut it :)
One of the factors that Hoegh-Guldberg and his ilk throw out with false certainty, is that ocean acidification is negatively affecting coral. But the emerging evidence is that coral are currently dealing with large swings in pH that are far greater than climate models project in the future.
Much of the acidification worries are due to the false assumption that ocean and atmospheric pCO2 is in equilibrium but rarely is that true.
As discussed in another essay pH drops dramatically with depth and in upwelling regions CO2/ DIC are brought the surface lowering pH while ventilating CO2 to the air. The biological pump then absorbs that upwelled CO2 via photosynthesis , raises the pH higher than what equilibrium would predict, by pumping the CO2 back to depth in various organic compounds
http://landscapesandcycles.net/ocean-acidification-natural-cycles—uncertainties.html
Although coral live in low nutrient environments typically away from upwelling sites, Kline shows that pH also varies daily and seasonally due to changes in photosynthesis and that coral basically control the range of pH in the reef
Ocean oscillations also affect pH as Pelejero 2005 wrote
The Calcification Paradox
In addition to those biological processes and natural cycles that control ocean pH, the process of calcification adds a mole of CO2 and H+ for every mole of calcium carbonate produced, while pumping buffering alkalinity to depth.
NIght time dissolution of coral rubble enhanced by lower pH helps increase alkalinity and recycle calcium and carbonate ions
Furthermore most coral have evolved vesicles in which they can control the pH and the process calcification, revealing a much lower sensitivity to changes in environmental pH than what has been suggested due to climate change.
Also researchers initially argued that coral would suffer from lower calcification rates due to the reduction of carbonate ions from lower pH. But what the new research is showing is coral do not absorb carbonate ions, they absorb bicarbonate ions which will increase with more CO2.
It is increasingly less certain that coral will be hurt by lower pH from CO2.And increasingly more certain that coral can increase their tolerance to thermal stress.
[In case you folk are actually interested in facts rather than just abusing each other.]
My site shows quantitatively that CO2 can cause no problem with ocean pH or coral calcification.
There are many eminent chemical and oceanographic journals who would be absolutely fascinated by your result. I look forward to seeing it in one of them.
So, you haven’t bothered to look?
[Just like these journals, they do not want non-AGW. I gave some examples of this in my site, which you won’t read – and by so doing, give a substantiating example of what I am stating. Thank you! ]
Jim Steele,
[posted out of sequence for scroll]
Except it isn’t evidence of my lack of understanding. The mechanism described by the Adaptive Bleaching Hypothesis is not the most intuitive fit for the classic definition of an evolutionary adaptation:
Daphne G. Fautin describes why she and Buddemeier chose the word “adaptive” over other possible alternatives:
Not being an expert, I can perhaps afford to not be so particular about terminology, and I’m not so long as everyone understands how terms are being used. Your descriptor “pre-adaptation” did immediately bring to mind the so-called industrial melanism of peppered moth fame — the darker phenotype was already extant but quite rare, thus arguably “pre-adapted” to tree barks increasingly darkened by sooty pollution as the Industrial Revolution progressed. Had not tree barks become darkened by coal soot, there would have been no driver for the shift in frequency to the “dark” allele amongst peppered moth populations, which is consistent with punctuated equilibrium. Not to put too fine a point on it, the rapidity of the change was almost certainly helped by the fact that a moth which is heterozygous dark/light is will have the dark phenotype because the dark allele is dominant.
Where the analogy does break is that the genotype of corals shifting/shuffling variants symbionts doesn’t change. Thus an individual coral polyp can “adapt” within the course of its own lifetime — “adapt” in “scare quotes” because the change in symbiotic partner is not genetically heritable. Regardless, that the shift/shuffle *can* occur within the lifetime of one individual is a clearly arguable advantage.
That said, my allegedly “unfounded snarkiness” mainly comes from the fact that you persistently ignore what *else* ABH proponents say about it in an over-confident manner — “pre-adapted” being yet another example of you peddling the notion that coral survivability under a regime of rapid climate change is a lock. From the same citation as above, again Dr. Fautin:
*Every* paper I’ve read that is supportive of ABH, including Fautin and Buddemeier’s seminal paper proposing it, includes caveats along the lines of the sentence bolded just above.
Except for Fautin and Buddemeier’s obvious irritation with Hoegh-Guldberg for his long-running arguments against ABH, neither they nor anyone else I know of in the coral research community has a tendency to throw him under the bus like you do — they tend to agree with him more often than not on most everything else, especially as seen by how frequently they cite his many works.
Until you face up to the fact that your virtual guarantees aren’t consistent with what supportive literature says about ABH, you’re pretty much going to be stuck complaining about me being an “Internet sniper” who is largely ignorant of corals. As I have freely volunteered several times before, I’m not even a working biologist, much less a corals expert. But my personal expertise here just isn’t relevant — I can read plain English and clearly see that what you say about literature in support of your arguments very frequently does not match what the literature itself actually says.
brandonrgates: This seems to be substantiated in patterns of “strains” of zooxanthellae that live in shaded and lighted portions of a single coral colony, and of “strains” of zooxanthellae that live in shallow and deep colonies of a single species of coral. Part of the decision on whether to use the pigeon-hole “adaptation” or “acclimation” that Hoegh-Guldberg raises may depend on one’s concept of who is “in charge” in the symbiosis – if the animal is making a selection, it may be nearer the “acclimation” end, whereas if the zooxanthella is choosing a suitable home, it may be nearer the “adaptation” end.
Despite the language of those experts, it is not an either/or. Each strain of zooxanthellae exhibits great variation in their progeny, and each strain of coral exhibits great variation in their progeny. In each interval of time of multiple generations, the strains with the most favorable survival in each new slightly modified environment are selected together.
I use the phrase “great variation” merely to stand for the fact that there is more random variability than has so far been measured and documented. That isn’t a revolutionary claim, merely an induction from all the other instances of adaptation to “relatively rapid” change in antibiotics in the bacterial environments, herbicides in the farm environments, DDT in the mosquito environments, and so on.
We also explicitly stated that the ABH applies to the level of bleaching under which the symbiosis evolved — what has been considered “background” – and that a mechanism that evolved under that level may not be adaptive if what we are now experiencing is as unprecedentedly severe and widespread as some believe (which is consistent with what Hoegh-Guldberg reported has been found in Japan).
Every mechanism in every strain so far studied displays variability in the progeny of every generation. (The people who claim “irreducible complexity of mechanism” are generally the “Intelligent Design” proponents, and they are always wrong.) The idea of a mechanism that has evolved in a level and may not be adaptive in a slightly chantged level is not what random variation and natural selection are about. The “mechanism” that has “evolved” has variation in every one of its components, and as the environment changes the relative abundance of the variations that survive to reproduction changes.
It may well be the case that you read and understand English perfectly, but the passages that you quote are not accurate representations of the great amounts of variability in the natural population. And even though the quotes are from experts in the field, it is still the case that those quotes misrepresent the role of natural variation and selection in evolution. Random variation is always underestimated. Each new medicine, herbicide, insecticide, or other small environmental change produces a noticeable effect for a while, and the target population is always resurgent. The most reasonable inference in this case is that the coral, their hosts, preditors, food populations, etc will all adapt, and produce new distributions of all the alleles relevant to all the mechanisms..
matthewrmarler,
No, that would just be the most comforting one. The more reasonable inference is the one supported by quantified estimates, not appeals to what every researcher in the field learns their first year of undergraduate coursework.
No, that would just be the most comforting one.
And you are taking an unreasonably alarming one.
1. Are there fewer species? Don’t know..
Studies always focus on macroscopic species. There are 100+ times more bacteria (prokaryotes) by mass or number of species than anything else.
Unless you have accurate data on Prokaryotes the claim there are fewer species is an unresearched talking point.
2. Are there fewer large species? Sure. Man has invaded all kinds of ecologies and modified or replaced them. Cities and farms displace native species. Commercial fishing has modified ocean populations. Species that have large territorial needs haven’t faired well.
3. Do we need all the species? No. 99% of all species are extinct. Obviously we didn’t need most of them. It makes no sense to save a snail darter. There are all sorts of local specializations where a parent species evolved to fill a local niche. There are some bird species that have a left and right coast population that can breed with the flyover country population but not each other. We don’t need all of them. It makes sense to keep some wolves around since they are the most diverse breed of dog, but we don’t need every single regional variant. We don’t need to preserve the Chihuahua. We can always crossbreed a wolf and a rat and recreate the species.
We would like to identify and preserve the parent species to the extent possible that diversified to fill the niches. After all as long as there is a cat- like animal you will get a saber toothed tiger when needed, it has happened at least half a dozen times independently.
I don’t feel too bad about the situation. The greens have destroyed more species with their biofuel initiatives and the rainforest destruction it led to than probably anyone else. They have as much or more blood on their hands than I do. The fact they are so fond of animal-killing windmills illustrates their insincerity.
The greens aren’t really serious about this species thing and it is just a talking point to push their policies.
4. Is CO2 a problem? No, definitely no.
More CO2 has created a more abundant ecosystem and saved far more species than it harmed. Blaming CO2 for species problems is somewhere between laughable and deluded.
But the species issue is a problem that should be looked at by more serious people (than greenies) and some choices made. The increasing global population is going to have to be accommodated.
Brandorgates,
You’re making a great deal of sense. That may be scant comfort when confronted with a sea of ignorance but you’re doing pretty well.
Anthony Purcell: “Brandorgates,
You’re making a great deal of sense. That may be scant comfort when confronted with a sea of ignorance but you’re doing pretty well.”
Awesome!
Tell us, when you’re not
playing with your Xboxmodelling sea level rise, do you moonlight as a stand-up comedian?brandonrgates: The more reasonable inference is the one supported by quantified estimates,
You are mixing things up again. There is little in that link relating coral loss to temperature or CO2 change. Not a single extinction has been attributed to the change in CO2 from 280 ppm to 400 ppm, or the ca 1C global mean temp increase over the last 125 years. And as I wrote, among the species not extinct are those that have been explicitly targeted by humans using chemical methods: bacteria, weeds, insects, viruses, spirochetes, etc. Beneficiaries of the increase in global mean temp and CO2 include the famous (now) coccolithophores and forests.
what every researcher in the field learns their first year of undergraduate coursework.
You have been writing about “adaptation” in the informal, non-Darwinian sense that ignores the random variation and natural selection. Regardless of what “everyone” learns, the experts whom you have quoted about corals and temp has underestimated the genetic random variation of the progeny in all of the species in every generation.
Anthony Purcell: That may be scant comfort when confronted with a sea of ignorance but you’re doing pretty well.
Not so. He skips around and has kept changing his points as scientific knowledge contradicting the extreme claims of the alarmists has been cited. He even wrote himself (that is, he presented a long quote) that he was referring to the common-sensical notion of adaptation, not the Darwinian sense of evolution via random variation and natural selection.
Consider his last link to an article, referencing extinctions not due to a 1C increase in global mean temp or the increase in CO2 from 280 ppm to 400 ppm. What large population of small rapidly-reproducing organisms has become extinct due to global climate change? Nothing as dramatic as the near total loss of American chestnut trees due to a naturally occurring virus in the early 20th century.
Brandin R Gates illustrates the difference between writing a whole lot of loosely related stuff and addressing a specific topic.
oops, Brandon, not Brandin. Sorry.
Anthony Purcell,
Thanks. I’ve enjoyed your comments on this thread as well. My favorite so far: To suggest that AIDS is not a problem because septicemic plague is worse would be equally fatuous.
It’s the selective ignorance that bugs me most, followed by the pretended knowledge and expertise. I suppose what I find most galling is the apparent lack of recognition that “uncertainty” means we’re all equally in the dark — something I see as the root of most evils in highly contentious partisan spats.
brgates, “I suppose what I find most galling is the apparent lack of recognition that “uncertainty” means we’re all equally in the dark — something I see as the root of most evils in highly contentious partisan spats.”
Bravo! One little hitch though, when you use ideal models and worst case scenarios you are skewing the uncertainty toward an unrealistic.probability. If we were all “equally” in the dark, you could do an average of the guesses, with a Bayesian approach of course, an ignore the partisan spats. Using that method the estimate of “sensitivity” is trending downward :)
matthewrmarler,
I know exactly what that paper says because I read it.
I’ve been quoting Daphne Fautin, who is the co-originator of ABH along with her husband Robert Buddemeier. You can quibble about *their* usage of “adaptation” all you like, but semantic arguments do not address the fact that ABH is rooted in the very mechanisms of variation and selection you mention.
That’s an unfounded comparison, Matthew. Neither you nor the quotes I’ve provided quantify the actual level of diversity. You’re basing conclusions on quantification that rest on a naive assumption that the unspecified (unknown?) level of diversity is sufficient to render Fautin’s et al. caveats and cautions moot. Here is one such caution from Fautin which I’ve previously quoted:
Until you can demonstrate with evidence that she’s wrong, you handwaving away her expert opinion with implicit presumption that you have a better handle on evolutionary and adaptive mechanisms than she does will remain an uncompelling argument.
What I’m illustrating is the process of taking as comprehensive view of the entire system as possible within the constraints time, space and my inexpert level of understanding the salient issues. What that means is that not every reference I provide will necessarily deal strictly with corals. No one paper has all the answers, and this is confounded by the fact that not all papers agree with each other. I have zero control over the complexity of the system, nor our incomplete and uncertain understanding of it. At best, I have control over my own learning curve … and let me tell you, it’s a steep one.
You might do well to stop faulting me for illustrating the reality of humanity’s collective ignorance. You would do particularly well to stop pretending you have better answers than domain experts who don’t reek of similar hubris when they go out of their way to describe what they know they do not know, and consider essential to figure out.
No worries. Call me “Brenda” and we’ll have words. :-)
brandonrgates: You might do well to stop faulting me for illustrating the reality of humanity’s collective ignorance.
That is absurd. I am addressing specific points. for example, there are no reports of (massive or isolated) extinctions of large populations of small rapidly growing organisms because of CO2 or 1C of warming; ABH is only one mechanism; the flaws in Marcott et al were serious; all attempts to specifically eradicate (or even diminish) pests by chemical means have produced adapted populations instead; people, including experts, frequently underestimate the amount of random variability in the systems that they study; corals and the populations that interact with them are selected together by the environment; other species have thrived with increased CO2 and temp, some ocean-dwelling. You are irrelevantly bringing in other factors besides CO2 and temp that are not the focus of the large attempt to eliminate fossil fuel use.
Taken all together, the details and broad points do not support a claim that there is a threat of coral extinction posed by CO2 and warming.
captdallas,
Thanks.
lol. We just got done agreeing that we’re all equally in the dark.
Several times I’ve mentioned that the Big Five extinctions wiped corals from the fossil record. Several times Steele has replied that I’m apparently unaware that modern corals come from an entirely separate lineage. Go ask him to apply an objective prior and finish his argument.
Lewis and Curry (2014) being one I presume:
All that really tells me is that choice of prior distribution affects the central estimate of the model output. Which is, like, academic.
It’s possible I need to read harder.
matthewrmarler,
It’s absurd to imply that I am not.
It has been said that those who do not learn history are doomed to repeat it:
And since it putatively allows individual corals to shift and shuffle within their own lifetimes, it is potentially the fastest.
Thanks for that reminder. I asked you for, and did not receive, an equivalent alternative which you find more acceptable.
The goal here is to not eradicate corals. What you keep glossing over is that nearly extirpating them probably wouldn’t be a good plan.
You also avoid like the plague (BTW, when was the last time plague was a catastrophic issue for us?):
1) evidence that corals *vanished* five — count them, one two three four five — FIVE times from the fossil record,
2) marine species were often amongst the most affected during those extinction events,
3) recovery from those events took on the order of hundreds of thousands to millions of years.
That probably doesn’t mean we necessarily face an existential crisis, but it doesn’t sound like easy living to me either.
You previously said always. You still haven’t quantified it. I assert you cannot because to do so would mean that the amount of actual variability is known. Considering that you haven’t actually demonstrated superior knowledge than the domain experts whom you’ve declared ignorant by fiat, you have essentially zero basis to conclude that allele diversity within a given species will adapt under any conceivable environmental/ecological change.
This is Biology 101 again. The implication that doctoral-level researchers in this domain do not understand what you claim to is presumption bordering on hubris.
Clearly some have, the fossil record does not show total gaps of all species due to any known mass extinction event.
Sorry, me pointing to literature suggesting that past ocean temperature and pH excursions have been lethal to most then-extant marine species is about as far from irrelevant in this context as I can imagine.
I don’t at all need to appeal to AGW/ocean acidification concerns to argue for *transitioning to* non-fossil sources of energy.
Only the ones you continue to insist we focus on. You bet the citations I’ve made are irrelevant to supporting your beliefs.
brgates, Actually it was Annan and Hargreaves, Bayesean methods tend to lop off fat tails just like Lewis and Curry. So if you consider the range to be about 0.5 to 4.0 C per doubling and RPC 4.5 to be somewhat realistic, we would be equally in the dark.
“Thanks for that reminder. I asked you for, and did not receive, an equivalent alternative which you find more acceptable.”
I take it you don’t approve of Rosenthal et al. who politely poked Marcott and Mann :)
captdallas,
How would I have a clue whether RCP4.5 is any more realistic than RCP6.0 or RCP8.5? My understanding is that we’re tracking closer to RCP4.5 right now, but I find no guarantee that will hold out to 2100.
More annoyed at Marler for repeating “Marcott is flawed” without providing an equitable alternative to support his claim. Since Rosenthal isn’t a global or even hemispheric reconstruction, it doesn’t much help. On the lighter side, I was amused that Dr. Curry sorta kinda implied that Mann’s critiques of Rosenthal was some kind of endorsement of the latter’s veracity.
Do we like Moberg (2005) in this forum?
http://www.ncdc.noaa.gov/paleo/globalwarming/images/moberg-2005-large.jpg
I’m guessing not:
https://upload.wikimedia.org/wikipedia/en/d/dd/IPPC_1990_MBH_1999_Moberg_2005.png
Besides, it doesn’t cover the entire Holocene, and is limited to the northern hemisphere.
You know, one reason to not like IPCC 1990 Figure 7.1.c is that it implies a higher CS than MBH99 and (possibly) Moberg (2005).
brgates, “Do we like Moberg (2005) in this forum?”
Not really though I thought it was a much better effort than Mann et al.
The interesting thing about “global” is that most focus on land based proxies which are inherently noisy, but their confidence intervals don’t reflect that. When a land based paleo reconstruction supposedly has better accuracy i.e. tighter error bars than instrumental my BS detector tends to sound :) Ocean paleo has the advantage of 1000 times greater heat capacity which helps compensate for the lower resolution. 50 year resolution is more climate relevant than annual that tends to actually be a few growing season months per year, annually. Since current warming is more winter time in the high northern latitude you are not comparing apples to apples are you?
brgates, “You know, one reason to not like IPCC 1990 Figure 7.1.c is that it implies a higher CS than MBH99 and (possibly) Moberg (2005).”
Odd thing about that implied higher sensitivity is that climate appears to be more sensitive to combined solar and volcanic forcing on longer time scales. That solar variability though is based on 10Be which doesn’t appear to be the same as plain old TSI. Kind of like the recent discovery of ~200 year variability of a couple degrees in the Himalayan ice cores related to the 205 year solar cycle which of course cannot have any impact right? I am inclined to believe something was missed in the theory.
captdallas,
I would naively assume that six years would yield some improvements.
Gonna need an example or two for that one … doesn’t fit my recollection.
As it should.
I more or less agree save for an SST proxy probably isn’t responding to a signal representing the entire oceans to depth.
I agree there. If you’ll pardon the pun, one perennial debate is about how to splice that to the instrumental record in a meaningful and “unbiased” way.
In a perfect world, I’d have a reconstruction of SSTs sampled from all the major reef systems across as broad a variety of latitudes as possible, while also taking care to distinguish between areas closer to eastern or western shores. But then there’s always the problem of individual locations being noisy, if not giving spurious signals … that is if I properly understand the issues.
Sure. I get really nifty fits to the instrumental record if I smooth out TSI with a trailing 132-month running mean. Problem is the regression coefficient doesn’t look at all physically plausible. Better analyses have been done with better methods than linear regressions and SMAs I’m sure.
I’ve not had much luck working volcanism in with TSI over paleo data.
Zero impact wouldn’t be my default assumption.
Shocker. :-)
Pingback: Weekly Climate and Energy News Roundup #228 | Watts Up With That?
https://www.yahoo.com/news/polygamy-could-save-great-barrier-000000626.html?nhp=1
“…a crucial discovery because it opens the door for adult coral to bind with algae that is better suited to protect it from the mass coral bleaching episodes that have recently rocked reefs across the globe.”
Currently there is concern about coral bleaching. I read a short interview with Valerie Taylor (Magazine, Weekend Australian, May 14). You may recall she made many documentaries with her husband Ron about sharks. She’s now 80, and still dives. She says,
“In 1965 we went from one end of the reef to the other, over six months, and we found bleaching then. In the ‘70s we went back and you’d never know it happened. The coral had recovered; nature had taken care of it. I’ve seen reefs in PNG that were as white as snow and I’ve just come back from there and they’re terrific.”
She thus shows that bleaching is reversible. Presumably spores from unaffected corals can flow in again to recolonise affected areas.
The current storms flowing down the East coast result from extensive seismic activity along Vanuatu’s coasts (around -14.25, 166.6) beginning about 06Apr16 (Mag7) and reaching zero km depth about 12Apr16, and continuing almost continually through to 30Apr16. I’m assuming the seismic accompanies lava. Vanuatu is on an anti-clockwise current upstream from Qld. Timing to reach Qld is about correct.
Bleaching probably results from upstream H2S from volcanic activity mentioned above, not CO2 (heating or acidity) or Man’s pollution; the northern areas where there’s little run-off are the ones affected. I expect there will be more bleaching from the cause of the current Eastern storms.
Brandon Gates,
Your skill is word smithing and redirecting a discussion. You and slandering sou are professional skeptic bashers. Criticisms that move the science forward are welcome but as can be seen by your posts you intent is to discredit any skeptic argument with dishnest comments
The only criticism you can mustered is that my comment of a “virtual” guarantee that coral can adapt to a 2C climate warming is that researchers have expressed greater uncertainty. True. But such disagreements are simply how science works. You can repeat that disagreement ad nauseum but you still fail to refute any of the evidence that support my arguments. Furthermore many of your quote-mining remarks regards uncertainty were from earlier papers before identification of heat tolerant symbionts.
My “virtual guarantee” is that bleaching and climate warming will not cause coral to go extinct . As I have pointed out from the very beginning, coral mortality due to bleaching is minor relative to other factors like storms. You dishonestly claimed I ignored those other factors, trying to redirect the discussion. Many of the researchers that expressed uncertainty is based on their uncertainty about climate model projections, projections whichthey lack any epertise to fully evaluate. On the other hand everything I have written refutes Hoegh-Guldberg’s ludicrous claim that we will lose 95% of our coral by mid century. Yet you never attack his outrageous claims and try to defend it with irrelevant remarks about a 6th extinction or by suggesting that his papers are cited that provides some sort of proof his projections are valid
My comment that there is a “virtual guarantee” is based on evidence that you have failed to refute. .
1. Coral thrived in tropical temperatures that were 2.1C warmer than today That strongly suggests coral can likewise survive any projected warming by climate models and that ocean temperatures that are now 2.1C cooler are not deadly. You have provided no evidence whatsoever that this is not the case.
2. New genetic techniques have identified symbionts that are pre-adapted to thermal stress and have allowed observations of coral rapidly acquiring these new symbionts. First you ignorantly mocked the idea of pre-adapted symbionts and rapid adaptation. As you checked out my citations you realized the evidence was against you.
3. Much of the reason for the expressed uncertainty by other researchers is we are just learning how symbiont shuffling and shifting works. Nonetheless the evidence shows coral are indeed rapidly adapting. Reefs that bleach recover within less than ten years. T
4. Much of the reason for the expressed uncertainty by other researchers is that they do not know which modeled projections to trust. You tried to promote a 6th extinction due to climate change, But the researcher you quote reports tropical ocean temperatures were capped at 31C (shallow seas like the Persian Gulf are slightly warmer) during the K/T. This limited increase in temperature coincided with estimates that CO2 levels were 5 to 10 times greater than today.
5. Could other human factors lead to coral extinction. Possibly, but that was not the point of the essay or the gurantee. It was just you way of trying to redirect the topic of the essay. You quote mine in order to juxtapose my one comment to a researcher’s uncertainty, yet you fail to acknowledge those researchers confirm most every point made in the essay.
6. As research has expanded there is a growing sense that coral are very resilient, and on the verge of a 6th extinction that you suggest. To repeat Dr. Smith observations of reefs like those of the Palmyra Atoll that are around uninhabited islands the reef look “almost exactly the same as it did 2,000 years ago. Far from any local human population, Palmyra’s reefs retain 80 to 90 percent of corals, despite past bleaching and ocean acidification.”
7. If instead of sniping at skeptics, if you sincerely want to help the coral then join me in advocating improving landscapes effects that are the real problem. Hoegh-Guldberg’s ludicrous claim that we will lose 95% of our coral by mid century is the only false certainty that we should be concerned with as it redirects our attention from the treal problem, problems we can fix.
Jim, before you proceed any further with the clown dancer Gates, I suggest you take a look at this.
http://libertygibbert.wordpress.com/2010/08/09/dobson-dykes-and-diverse-disputes/
catweazle666, Indeed, Gates is the perfect example of a post modern mamba
http://libertygibbert.wordpress.com/2010/08/09/dobson-dykes-and-diverse-disputes/
Jim Steele,
When the main topic of one of your essays is about what we might do to mitigate the effects of overfishing, dynamite fishing, pollution runoff, etc., I will happily advocate with you for that sort of reef preservation effort.
Regardless of the topic of any of your essays, I will myself be openly skeptical of over-confident claims, e.g. virtual guarantees, which are not supported by your own citations.
Brandon Gates says
Ahhhhh but my citations supported everything in the essay. And you failed to address the first 6 points that refute a 6th extinction that you and Hoegh-Guldberg are peddling!
https://wattsupwiththat.com/2016/06/03/expert-scientists-exaggerated-coral-bleaching-story/